Cricetids (Rodentia, Mammalia) from the Late Miocene Yihachi locality of Gansu, China
QIU Zhu-Ding, Lawrence J. FLYNN, WANG Ban-Yue, LI Lu
Vertebrata Palasiatica
2026, 64 ( 1):
1-25.
DOI: 10.19615/j.cnki.2096-9899.251117
Our purpose in this paper is to describe the hamster-like rodents (Cricetidae) from a Late Miocene age site in Linxia Basin, Gansu Province, and discuss their significance for the changing ecology of central Asia. The micromammal site known as Yihachi was introduced previously (Qiu et al., 2023; Qiu and Li, 2023), when its squirrels were discussed in some detail. We take this opportunity to describe the more abundant cricetids. There are four genera, common Nannocricetus and Sinocricetus plus the less abundant living Mesocricetus. A few specimens represent the high-crowned and lophodont Rhinocerodon. The cricetids and other faunal elements indicate an early Late Miocene age, and the pattern of occurrence of the hamster species is consistent with a picture of a changing paleoenvironment due to increasing effects of the East Asia monsoon system. After the disappearance of older and archaic genera, Yihachi represents growing endemism in the Late Miocene of northern China due to increasing seasonal rain and the declining average temperature.
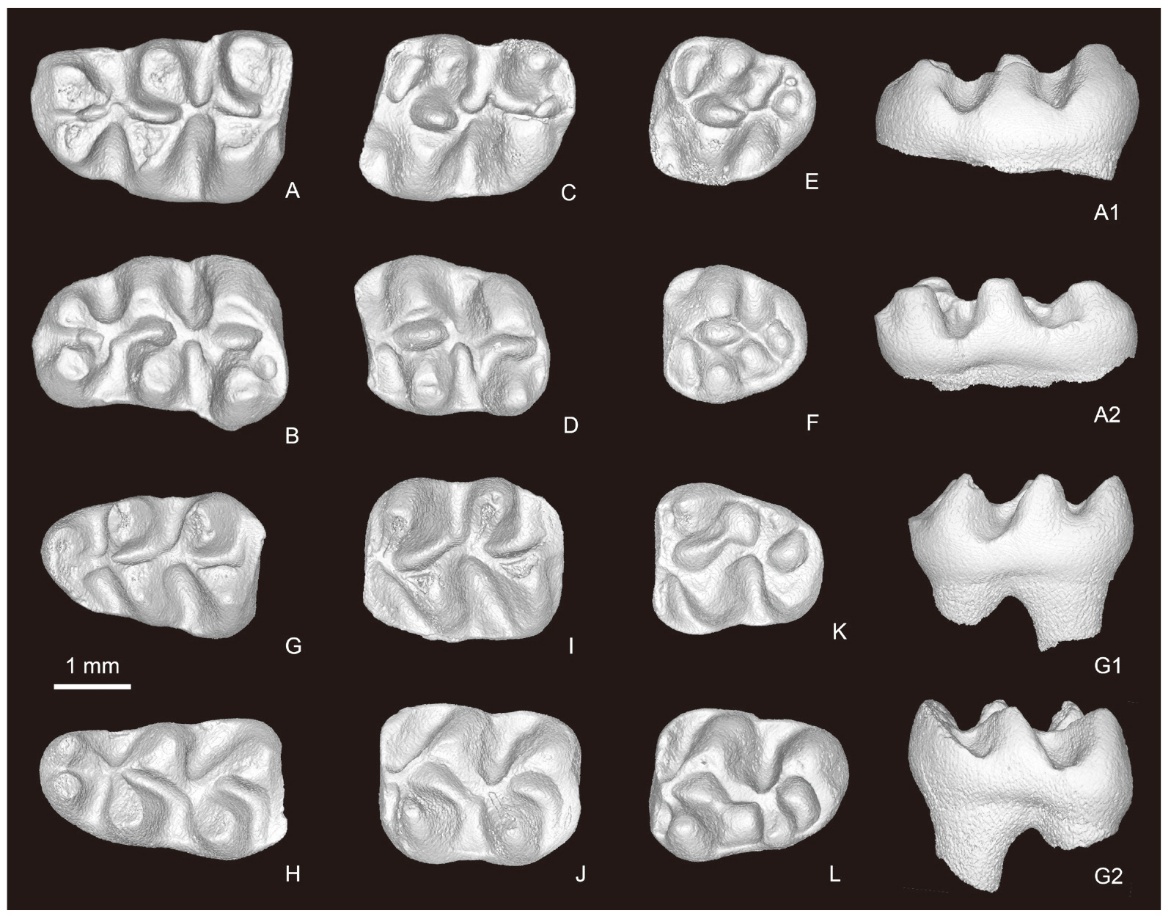
Fig. 2
Molars of Nannocricetus primitivus from Yihachi, Gansu
A. l M1 (IVPP V34185.376); B. r M1 (V34185.1); C. l M2 (V34185.377); D. r M2 (V34185.2);
E. l M3 (V34185.378); F. r M3 (V34185.3); G. l m1 (V34185.379); H. r m1 (V34185.4); I. l m2 (V34185.380);
J. r m2 (V34185.5); K. l m3 (V34185.381); L. r m3 (V34185.6)
A-L. occlusal views; A1, G1. lingual views; A2, G2. buccal views
Extracts from the Article
(Figs. 2-4; Table 1)
The M1 (Fig. 2A, B) is reniform in outline, with a slightly narrower anterior part. The anterocone is wide and bifid anteriorly. The lingual and buccal anterocones are nearly equal in size, both equipped with a posterior spur that joins together into the anterolophule posteriorly. The anterolophule is low, joining the protocone posteriorly. The protoloph I is absent in 102 of 106 specimens, and blurred in the others. The protoloph II is always present in the available specimens, though short and low. Most specimens lack a mesoloph, but a very short or remnant mesoloph is present in 12 of the observed specimens. The entoloph is marked, but short, connecting the hypocone to the yoke of the protocone posterior arm and the protoloph II. The posteroloph extends to the posterior base of the metacone. Among the 15 specimens from the lower lens (L-YHC) that can be observed, 14 have 3 roots and only one has 4 roots, while of 46 specimens from the upper lens (U-YHC) only 4 have 3 roots but 42 are 4-rooted. The M2 (Fig. 2C, D) is subrectangular slightly wider anteriorly than posteriorly. The anteroloph is prominent, extending to the lateral sides of protocone and paracone, respectively. There is a short but prominent anterolophule, connecting the middle of anteroloph and the junction of the anterior arm of the protocone and the protoloph I. Both the protoloph I and II exist, though the protoloph II is less developed. A short mesoloph can be seen in 3 or 4 cases in the teeth either from U-YHC and L-YHC, and remnant mesolophs touching the metacone are frequently present in the samples. The entoloph is short, connecting the hypocone to the yoke of the posterior arm of the protocone and the protoloph II. The posteroloph is developed, joining the posterior base of the metacone. Most specimens have 4 roots (2 lingual and 2 buccal roots), and a few are 3-rooted (one anteroposteriorly-elongated lingual root sometimes with a vertical groove along the lingual side of the tooth, plus 2 buccal roots). There are more 3-rooted M2s in the lower lens than in the upper. The M3 (Fig. 2E, F) is subtriangular in shape. It has similar structures in the anterior portion as in M2. The hypocone and metacone are reduced to a ridge-like cuspule, enclosing a small pit posteriorly with the posteroloph. There are 3 roots.
The m1 (Fig. 2G, H) is elongated and tapers anteriorly. The anteroconid is narrow and equipped with a distinct anterolophid. It is slightly bifid in most specimens, and usually has a cleft at its apex in fresh teeth. The anterolophulid is low, joining the anteroconid to the junction of the metalophid I and the anterior arm of the protoconid. The metalophid I is developed in all cases, but the metalophid II is lacking. There is no sign of a mesolophid. The hypolophid is poorly developed, anteriorly directed, and joins the ectolophid. The posterolophid is relatively strong, running from the hypoconid to the postero-lingual corner of the tooth but failing to enclose the posterosinusid. The ectolophid is curved, connecting the hypoconid and the protoconid. There are two roots. The m2 (Fig. 2I, J) is rectangular in occlusal surface. It is very similar to the m1 in general morphology, except in lacking an anteroconid and having a wider anterolophid buccally than lingually. In addition, a low but distinct mesolophid can be observed in 3 specimens from L-YHC and 2 from U-YHC. The m3 (Fig. 2K, L) is subtriangular, tapering posteriorly. It is like the m2 in morphology, but the hypoconid and entoconid are rather reduced, and the posterosinusid is enclosed.
Other Images/Table from this Article
-
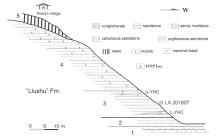 Fig. 1
Section of the fossil locality Yihachi
Fig. 1
Section of the fossil locality Yihachi
-
 Fig. 3
Size ranges and averages of length and width in the first molars of various species of Nannocricetus from Yihachi, Gansu and other localities in China Measurements of Lantian N. primitivus, N. wuae, Ertemte N. mongolicus and N. qiui are cited from Zhang et al., 2008, 2011; Wu, 1991, and Li et al., 2018, respectively Numbers inside the parentheses are specimen numbers
Fig. 3
Size ranges and averages of length and width in the first molars of various species of Nannocricetus from Yihachi, Gansu and other localities in China Measurements of Lantian N. primitivus, N. wuae, Ertemte N. mongolicus and N. qiui are cited from Zhang et al., 2008, 2011; Wu, 1991, and Li et al., 2018, respectively Numbers inside the parentheses are specimen numbers
-
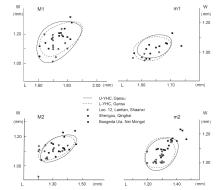 Fig. 4
Scatter diagrams showing the length and width of the first two molars of Nannocricetus primitivus from Yihachi, Gansu and some other localities in northern China Measurements of specimens from Lantian, Shengou and Baogeda Ula are cited from Zhang et al., 2008, Qiu and Li, 2008, 2016
Fig. 4
Scatter diagrams showing the length and width of the first two molars of Nannocricetus primitivus from Yihachi, Gansu and some other localities in northern China Measurements of specimens from Lantian, Shengou and Baogeda Ula are cited from Zhang et al., 2008, Qiu and Li, 2008, 2016
-
 Table 1
Measurements of molars of Nannocricetus primitivus from Yihachi, Gansu (mm)
Table 1
Measurements of molars of Nannocricetus primitivus from Yihachi, Gansu (mm)
-
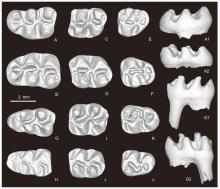 Fig. 5
Molars of Sinocricetus primus sp. nov. from Yihachi, Gansu A. l M1 (IVPP V34186, holotype); B. r M1 (V34188.1); C. l M2 (V34187.1); D. r M2 (V34188.2); E. l M3 (V34187.2); F. r M3 (V34188.3); G. l m1 (V34188.4); H. r m1 (V34187.3); I. l m2 (V34188.5); J. r m2 (V34187.4); K. l m3 (V34188.6); L. r m3 (V34187.5) A-L. occlusal views; A1, G1. lingual views; A2, G2. buccal views
Fig. 5
Molars of Sinocricetus primus sp. nov. from Yihachi, Gansu A. l M1 (IVPP V34186, holotype); B. r M1 (V34188.1); C. l M2 (V34187.1); D. r M2 (V34188.2); E. l M3 (V34187.2); F. r M3 (V34188.3); G. l m1 (V34188.4); H. r m1 (V34187.3); I. l m2 (V34188.5); J. r m2 (V34187.4); K. l m3 (V34188.6); L. r m3 (V34187.5) A-L. occlusal views; A1, G1. lingual views; A2, G2. buccal views
-
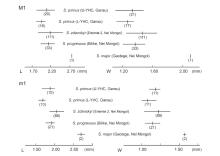 Fig. 6
Size ranges and averages of length and width in the first molars of Sinocricetus primus sp. nov. from Yihachi, Gansu and other species of the genus in China Measurements of S. zdanskyi from Ertemte, S. progressus from Bilike, and S. major from Gaotege are cited from Wu, 1991; Qiu and Storch, 2000; Li, 2010, respectively Numbers inside the parentheses are specimen numbers
Fig. 6
Size ranges and averages of length and width in the first molars of Sinocricetus primus sp. nov. from Yihachi, Gansu and other species of the genus in China Measurements of S. zdanskyi from Ertemte, S. progressus from Bilike, and S. major from Gaotege are cited from Wu, 1991; Qiu and Storch, 2000; Li, 2010, respectively Numbers inside the parentheses are specimen numbers
-
Table 2
Measurements of molars of Sinocricetus primus sp. nov. from Yihachi, Gansu (mm)
-
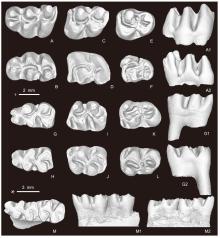 Fig. 7
Molars of Mesocricetus fengi sp. nov. from Yihachi, Gansu A. l M1 (IVPP V34189, holotype); B. r M1 (V34190.2); C. l M2 (V34190.3); D. l M2 (V34191.1, reversed); E. l M3 (V34190.4); F. r M3 (V34190.5); G. l m1 (V34190.6); H. r m1 (V34190.7); I. l m2 (V34190.8); J. r m2 (V34190.9); K. l m3 (V34190.10); L. r m3 (V34190.11), M. right mandibular fragment with m1-2 (V34190.1) A-M. occlusal views; A1, G1, M1. lingual views; A2, G2, M2. buccal views. Scale bars: + for A-L, * for M
Fig. 7
Molars of Mesocricetus fengi sp. nov. from Yihachi, Gansu A. l M1 (IVPP V34189, holotype); B. r M1 (V34190.2); C. l M2 (V34190.3); D. l M2 (V34191.1, reversed); E. l M3 (V34190.4); F. r M3 (V34190.5); G. l m1 (V34190.6); H. r m1 (V34190.7); I. l m2 (V34190.8); J. r m2 (V34190.9); K. l m3 (V34190.10); L. r m3 (V34190.11), M. right mandibular fragment with m1-2 (V34190.1) A-M. occlusal views; A1, G1, M1. lingual views; A2, G2, M2. buccal views. Scale bars: + for A-L, * for M
-
Table 3
Measurements of molars of Mesocricetus fengi sp. nov. from Yihachi, Gansu (mm)
-
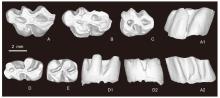 Fig. 8
Molars of Rhinocerodon linxiamys sp. nov. from Yihachi, Gansu A. r M1 (IVPP V34192, holotype, reversed); B. r M1 (V34193.1, reversed); C. l M3 (V34194.1); D. l m1 (V34193.2); E. l m2 (V34194.2). A-E. occlusal views; A1, D1. lingual views; A2, D2. buccal views
Fig. 8
Molars of Rhinocerodon linxiamys sp. nov. from Yihachi, Gansu A. r M1 (IVPP V34192, holotype, reversed); B. r M1 (V34193.1, reversed); C. l M3 (V34194.1); D. l m1 (V34193.2); E. l m2 (V34194.2). A-E. occlusal views; A1, D1. lingual views; A2, D2. buccal views
-
 Fig. 9
Scatter diagrams showing length and width in the first molars of Rhinocerodon from Yihachi, Gansu and other species of the genus from Kazakhstan and Nei Mongol Measurements of the specimens of the Kazakhstan species are cited from Zazhigin, 2003
Fig. 9
Scatter diagrams showing length and width in the first molars of Rhinocerodon from Yihachi, Gansu and other species of the genus from Kazakhstan and Nei Mongol Measurements of the specimens of the Kazakhstan species are cited from Zazhigin, 2003
-
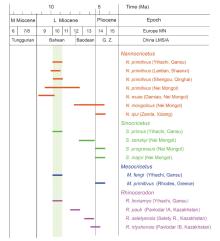 Fig. 10
Biostratigraphic ranges of various species of the cricetid genera present in the Yihachi Fauna G. Z. in the China LMS/A column indicates Gaozhuangian; Shading indicates the proposed ages of Yihachi cricetid composition
Fig. 10
Biostratigraphic ranges of various species of the cricetid genera present in the Yihachi Fauna G. Z. in the China LMS/A column indicates Gaozhuangian; Shading indicates the proposed ages of Yihachi cricetid composition
|

{kind=link}