
Vertebrata Palasiatica ›› 2025, Vol. 63 ›› Issue (4): 277-322.DOI: 10.19615/j.cnki.2096-9899.250912
Previous Articles Next Articles
LIU Jin-Yi1( ), Zhijie Jack TSENG2,3,4, JIN Chang-Zhu1, ZHENG Long-Ting5, SUN Cheng-Kai6, TONG Hao-Wen1,7
), Zhijie Jack TSENG2,3,4, JIN Chang-Zhu1, ZHENG Long-Ting5, SUN Cheng-Kai6, TONG Hao-Wen1,7
Received:2025-07-04
Online:2025-10-20
Published:2025-10-28
Contact:
liujinyi@ivpp.ac.cn
刘金毅1(), 曾志杰2,3,4, 金昌柱1, 郑龙亭5, 孙承凯6, 同号文1,7
基金资助:CLC Number:
LIU Jin-Yi, Zhijie Jack TSENG, JIN Chang-Zhu, ZHENG Long-Ting, SUN Cheng-Kai, TONG Hao-Wen. Eco-functional divergence of Crocuta and Pachycrocuta: a Huainan specimen reappraisal. Vertebrata Palasiatica, 2025, 63(4): 277-322.
刘金毅, 曾志杰, 金昌柱, 郑龙亭, 孙承凯, 同号文. 2025, 63(4): 277-322, 斑鬣狗(Crocuta)和硕鬣狗(Pachycrocuta)的生态功能之分化——对淮南标本的重新评估. 古脊椎动物学报.
Add to citation manager EndNote|Ris|BibTeX
URL: https://www.vertpala.ac.cn/EN/10.19615/j.cnki.2096-9899.250912
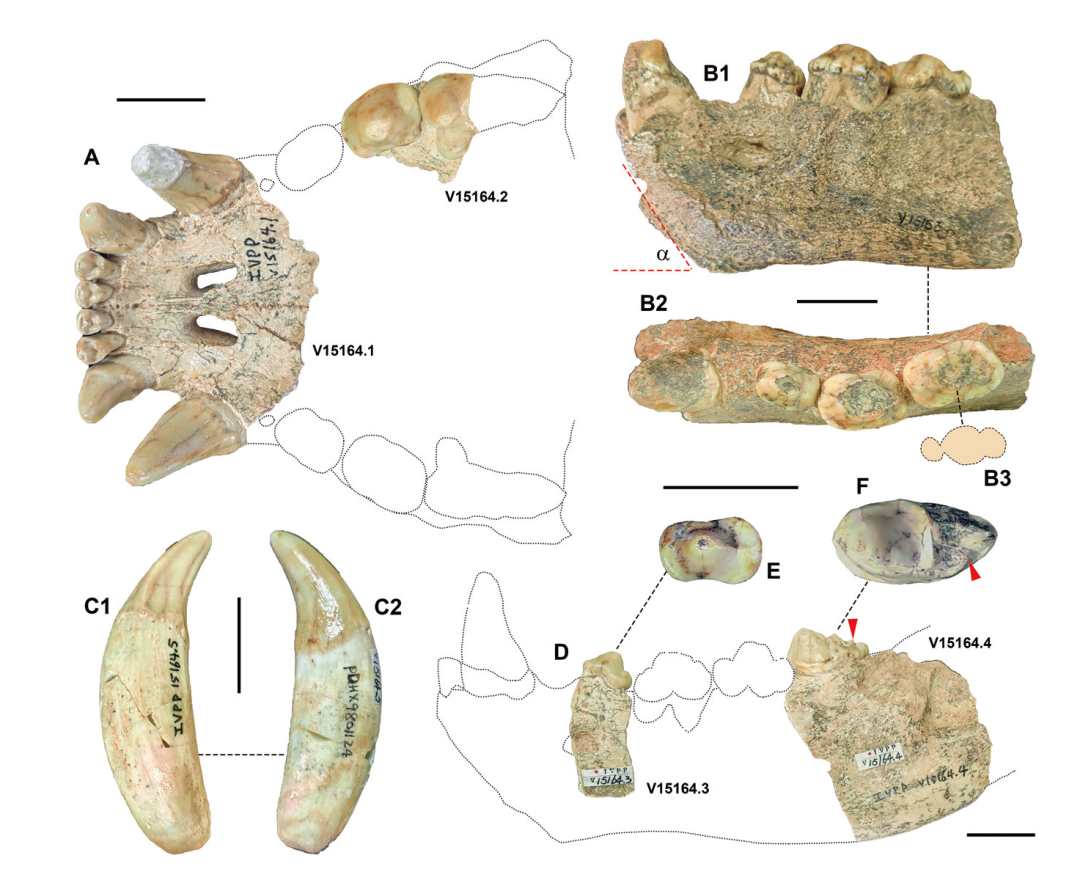
Fig. 1 The specimens of Pachycrocuta perrieri from Xiliexi of Dajushan, Huainan, Anhui, China A. a virtual maxillary outline in ventral view, showing IVPP V15164.1, V15164.2 and their specific position; B. a left fractured mandible V15163: B1. buccal view, B2. occlusal view, B3. an outline of the abrasive surface of p4; C. right lower canines: C1. V15164.5, in lingual view; C2. V15164.6, in labial view; D. a virtual mandibular outline in buccal view, showing V15164.3, V15164.4 and their specific position; E. enlarged crown view of p2 (V15164.3); F. enlarged crown view of m1 (V15164.4) The angle (α) in B1 is 54.50º. The red triangles in D and F point to the rear-most point of the trigonid Scale bars = 2 cm
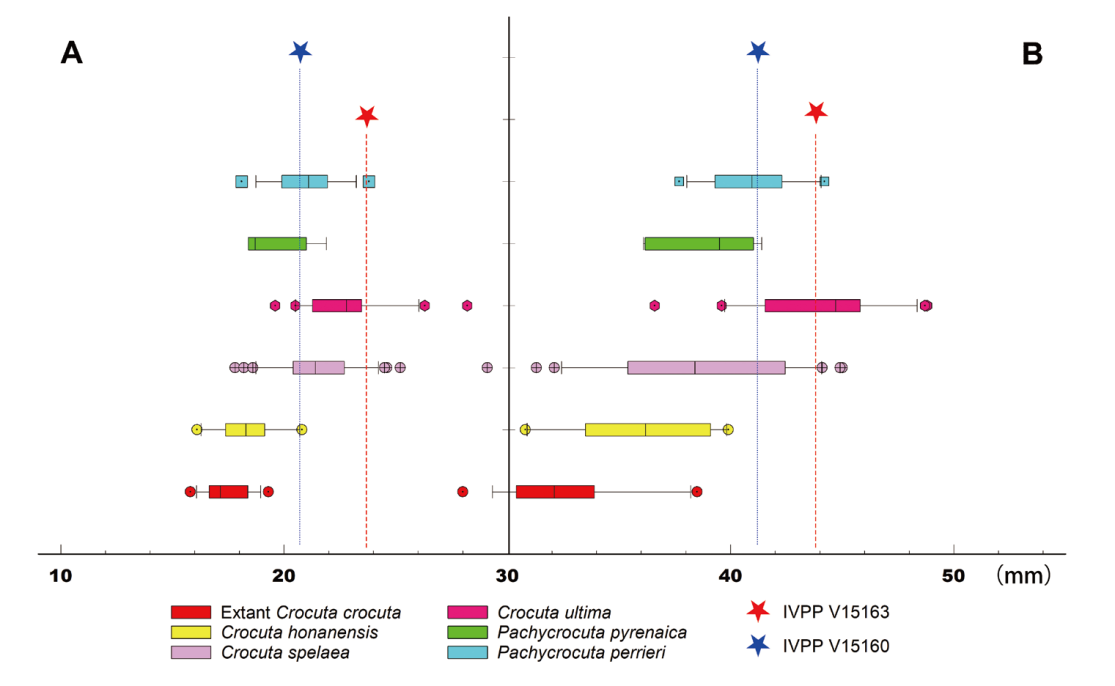
Fig. 2 Morphological comparison of the anterior end of the rostrum of hyenas in ventral view Left column: Crocuta type; Crocuta crocuta: A1 (IVPP OV232) and A2 (OV36); C. honanensis: A3 (IVPP V7294); C. ultima: A4 (LXD0007) Right column: Hyaena type; Pachycrocuta perrieri: B1 (IVPP V15164.1) and B3 (V27806); Parahyaena brunnea: B2 (AMNH M81834); Hyaena hyaena: B4 (AMNH M81228) L1 and L2 refer to the total length of the anterior palatine foramen and the palatine fissure, respectively Not scaled
| Huainan, Anhui | Crocuta | Pachycrocuta | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| XLX | DDS | crocuta | honanensis | spelaea | ultima | pyrenaica | perrieri | ||||||||||||||||||||||
| *3 | *4.1-5 | *4.6 | X | *0 | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | |
| c L | 18.7 | 16.9 | 17.4 | 17.67 | 32 | 13.4-19 | 15.47 | 1.28 | 19 | 12.4-15.1 | 13.94 | 0.78 | 52 | 14.9-19.8 | 16.80 | 1.02 | 33 | 15.2-22 | 17.47 | 1.56 | 14 | 15.1-18.24 | 17.05 | 0.90 | 21 | 15.8-19 | 17.58 | 0.71 | |
| c W | 15.1 | 14.1 | 14.3 | 14.5 | 35 | 11.2-13.6 | 12.23 | 0.58 | 18 | 10.1-13.5 | 11.68 | 0.92 | 52 | 12.5-15.8 | 13.68 | 0.70 | 32 | 12.6-15.7 | 14.31 | 0.80 | 14 | 11-14.9 | 12.90 | 0.99 | 21 | 13-16.3 | 14.26 | 0.73 | |
| p2 L | 15.7 | 15.3 | 15.5 | 16.1 | 187 | 12.3-18.1 | 14.78 | 0.98 | 19 | 12.4-15.3 | 13.83 | 0.76 | 434 | 13.2-19.3 | 16.70 | 0.94 | 40 | 15.1-18.8 | 16.92 | 0.90 | 16 | 14.3-17.4 | 15.64 | 0.75 | 48 | 13.3-17.7 | 15.43 | 1.05 | |
| p2 W | 11.1 | 9.9 | 10.5 | 11.1 | 76 | 8.2-12.7 | 10.33 | 0.92 | 19 | 8.3-10.5 | 9.49 | 0.54 | 383 | 10.4-14.4 | 12.40 | 0.80 | 39 | 10.1-14 | 12.26 | 0.95 | 16 | 9.4-10.4 | 9.80 | 0.30 | 47 | 9.1-12.9 | 10.55 | 0.80 | |
| p3 L | 21.8 | 21.8 | 22.7 | 190 | 17.5-23 | 20.15 | 0.96 | 22 | 16.2-19.2 | 18.24 | 0.76 | 528 | 19.6-26.5 | 22.57 | 1.01 | 49 | 21.2-26 | 23.24 | 1.11 | 21 | 15.7-21.5 | 20.11 | 1.15 | 52 | 19.2-22.9 | 20.78 | 0.76 | ||
| p3 W | 14.5 | 14.5 | 16.6 | 74 | 12.6-16.2 | 14.54 | 0.84 | 20 | 12.4-14.3 | 13.45 | 0.58 | 511 | 13.7-19 | 16.51 | 0.82 | 48 | 14.2-18.8 | 16.85 | 0.89 | 20 | 10.7-13.9 | 12.94 | 0.74 | 49 | 12.2-16.6 | 14.51 | 0.88 | ||
| p4 L | 23.9 | 23.9 | 24.3 | 177 | 19-24.6 | 21.97 | 0.97 | 22 | 19-21.9 | 20.68 | 0.77 | 528 | 21-26.8 | 24.19 | 1.01 | 48 | 20.4-26.4 | 24.28 | 1.14 | 23 | 19.7-24.4 | 22.51 | 1.09 | 56 | 21.4-25.2 | 23.14 | 0.99 | ||
| p4 W | 14.9 | 14.9 | 14 | 74 | 10.7-14 | 12.75 | 0.76 | 22 | 11.7-13.6 | 12.73 | 0.50 | 522 | 12.6-17.3 | 15.07 | 0.74 | 49 | 12.7-17.1 | 15.11 | 0.86 | 21 | 9.3-13.8 | 12.88 | 0.92 | 50 | 13.1-16.11 | 14.54 | 0.76 | ||
| m1 L | 24.5 | 24.5 | 176 | 23.3-31.2 | 27.26 | 1.71 | 20 | 23.1-26.5 | 24.91 | 0.91 | 442 | 27.5-38.4 | 32.45 | 1.46 | 47 | 29.7-37.4 | 32.77 | 1.71 | 23 | 22.7-26.3 | 24.33 | 0.99 | 60 | 23-27.6 | 25.26 | 1.26 | |||
| m1 W | 13.0 | 13 | 84 | 9.8-14 | 11.62 | 0.75 | 19 | 10.8-12.3 | 11.52 | 0.48 | 442 | 11.5-15.9 | 13.87 | 0.69 | 46 | 11.7-15.1 | 13.94 | 0.77 | 24 | 10.8-12.82 | 11.83 | 0.56 | 53 | 11-14.81 | 12.82 | 0.81 | |||
| m1tri L | 20.2 | 20.2 | 87 | 21.7-28.2 | 25.20 | 1.53 | 18 | 19.3-22 | 21.03 | 0.79 | 428 | 25.5-33.5 | 29.30 | 1.27 | 35 | 27.2-34.7 | 29.65 | 1.69 | 19 | 18.1-21.2 | 19.64 | 0.90 | 49 | 19-22.8 | 20.91 | 1.02 | |||
| p2 L /m1 L | 0.62 | 0.62 | 171 | 0.47-0.62 | 0.54 | 0.03 | 16 | 0.5-0.63 | 0.56 | 0.03 | 162 | 0.43-0.62 | 0.52 | 0.03 | 29 | 0.43-0.58 | 0.52 | 0.03 | 10 | 0.59-0.76 | 0.65 | 0.05 | 37 | 0.52-0.68 | 0.61 | 0.04 | |||
| p3 W /p3 L | 0.67 | 0.67 | 0.73 | 74 | 0.63-0.79 | 0.71 | 0.03 | 20 | 0.68-0.8 | 0.74 | 0.03 | 505 | 0.6-0.83 | 0.73 | 0.03 | 48 | 0.67-0.79 | 0.73 | 0.03 | 20 | 0.61-0.71 | 0.64 | 0.02 | 49 | 0.58-0.78 | 0.70 | 0.04 | ||
| p3 W /p4 W | 0.97 | 0.97 | 1.19 | 71 | 1.05-1.22 | 1.14 | 0.03 | 19 | 0.99-1.11 | 1.06 | 0.03 | 299 | 0.85-1.2 | 1.10 | 0.04 | 40 | 1.0-1.2 | 1.11 | 0.04 | 20 | 0.95-1.15 | 1.01 | 0.04 | 44 | 0.91-1.11 | 1.01 | 0.03 | ||
| m1tri L /m1 L | 0.82 | 0.82 | 84 | 0.88-0.95 | 0.91 | 0.02 | 17 | 0.81-0.88 | 0.85 | 0.02 | 422 | 0.86-0.95 | 0.90 | 0.02 | 35 | 0.85-0.96 | 0.90 | 0.02 | 18 | 0.76-0.83 | 0.80 | 0.02 | 49 | 0.74-0.87 | 0.83 | 0.02 | |||
| m1tri L /p4 L | 0.85 | 83 | 1.03-1.3 | 1.14 | 0.06 | 18 | 0.9-1.06 | 1.01 | 0.04 | 179 | 1.08-1.37 | 1.21 | 0.05 | 24 | 1.11-1.28 | 1.20 | 0.05 | 13 | 0.79-1.0 | 0.87 | 0.06 | 35 | 0.85-1.02 | 0.90 | 0.04 | ||||
| C L | 16.95 | 16.95 | 40 | 13.4-19 | 16.36 | 1.13 | 6 | 13-15.9 | 14.47 | 0.86 | 31 | 15.1-22.5 | 17.37 | 1.30 | 22 | 16-20.5 | 18.25 | 1.41 | 7 | 17-18.2 | 17.33 | 0.43 | 14 | 16.1-20.8 | 18.61 | 1.25 | |||
| C W | 13.1 | 13.1 | 41 | 10.2-14 | 11.87 | 0.81 | 6 | 10.9-11.9 | 11.35 | 0.33 | 30 | 11.7-15.7 | 13.46 | 0.98 | 22 | 12.5-15.2 | 13.92 | 0.69 | 7 | 12.4-13.1 | 12.66 | 0.24 | 11 | 12.4-15.5 | 13.65 | 0.84 | |||
| P3 L | 22.2 | 22.2 | 190 | 19-25.4 | 21.53 | 1.17 | 7 | 20-21.5 | 20.61 | 0.54 | 374 | 20.7-27.7 | 24.51 | 1.16 | 55 | 19.7-28.3 | 25.38 | 1.62 | 11 | 20.6-23.6 | 22.23 | 0.79 | 55 | 21-25.8 | 23.51 | 1.08 | |||
| P3 W | 16.8 | 16.8 | 80 | 14.2-18.7 | 16.56 | 0.99 | 7 | 14.1-16 | 15.17 | 0.57 | 360 | 15.4-20.5 | 18.21 | 0.91 | 55 | 15.8-21.7 | 19.05 | 1.34 | 8 | 13.73-15.9 | 15.09 | 0.67 | 46 | 14-18.8 | 16.58 | 1.05 | |||
| P4 W | 21.1 | 21.1 | 51 | 16.6-22.5 | 20.28 | 1.36 | 8 | 15-20.4 | 19.08 | 1.65 | 337 | 18-26 | 22.16 | 1.23 | 52 | 19.5-27.9 | 22.57 | 1.78 | 12 | 18.06-22.7 | 19.91 | 1.35 | 47 | 17.9-24.1 | 20.86 | 1.32 | |||
| P4ps L | 11 | 11 | 15 | 6.1-8.9 | 7.48 | 0.74 | 8 | 7.9-9.7 | 8.98 | 0.54 | 29 | 7.4-11.4 | 9.52 | 1.19 | 37 | 7-12.7 | 8.98 | 1.08 | 6 | 9.3-11.3 | 10.43 | 0.71 | 33 | 9.72-13.5 | 11.57 | 1.06 | |||
| P3 W /P3 L | 0.76 | 0.76 | 80 | 0.68-0.86 | 0.75 | 0.03 | 7 | 0.66-0.76 | 0.74 | 0.03 | 345 | 0.67-0.86 | 0.74 | 0.03 | 53 | 0.69-0.83 | 0.75 | 0.04 | 8 | 0.62-0.77 | 0.68 | 0.04 | 46 | 0.6-0.78 | 0.71 | 0.04 | |||
Table 1 The dental measurements and comparisons with other bone-cracking hyenas (mm)
| Huainan, Anhui | Crocuta | Pachycrocuta | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| XLX | DDS | crocuta | honanensis | spelaea | ultima | pyrenaica | perrieri | ||||||||||||||||||||||
| *3 | *4.1-5 | *4.6 | X | *0 | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | n | range | X | s | |
| c L | 18.7 | 16.9 | 17.4 | 17.67 | 32 | 13.4-19 | 15.47 | 1.28 | 19 | 12.4-15.1 | 13.94 | 0.78 | 52 | 14.9-19.8 | 16.80 | 1.02 | 33 | 15.2-22 | 17.47 | 1.56 | 14 | 15.1-18.24 | 17.05 | 0.90 | 21 | 15.8-19 | 17.58 | 0.71 | |
| c W | 15.1 | 14.1 | 14.3 | 14.5 | 35 | 11.2-13.6 | 12.23 | 0.58 | 18 | 10.1-13.5 | 11.68 | 0.92 | 52 | 12.5-15.8 | 13.68 | 0.70 | 32 | 12.6-15.7 | 14.31 | 0.80 | 14 | 11-14.9 | 12.90 | 0.99 | 21 | 13-16.3 | 14.26 | 0.73 | |
| p2 L | 15.7 | 15.3 | 15.5 | 16.1 | 187 | 12.3-18.1 | 14.78 | 0.98 | 19 | 12.4-15.3 | 13.83 | 0.76 | 434 | 13.2-19.3 | 16.70 | 0.94 | 40 | 15.1-18.8 | 16.92 | 0.90 | 16 | 14.3-17.4 | 15.64 | 0.75 | 48 | 13.3-17.7 | 15.43 | 1.05 | |
| p2 W | 11.1 | 9.9 | 10.5 | 11.1 | 76 | 8.2-12.7 | 10.33 | 0.92 | 19 | 8.3-10.5 | 9.49 | 0.54 | 383 | 10.4-14.4 | 12.40 | 0.80 | 39 | 10.1-14 | 12.26 | 0.95 | 16 | 9.4-10.4 | 9.80 | 0.30 | 47 | 9.1-12.9 | 10.55 | 0.80 | |
| p3 L | 21.8 | 21.8 | 22.7 | 190 | 17.5-23 | 20.15 | 0.96 | 22 | 16.2-19.2 | 18.24 | 0.76 | 528 | 19.6-26.5 | 22.57 | 1.01 | 49 | 21.2-26 | 23.24 | 1.11 | 21 | 15.7-21.5 | 20.11 | 1.15 | 52 | 19.2-22.9 | 20.78 | 0.76 | ||
| p3 W | 14.5 | 14.5 | 16.6 | 74 | 12.6-16.2 | 14.54 | 0.84 | 20 | 12.4-14.3 | 13.45 | 0.58 | 511 | 13.7-19 | 16.51 | 0.82 | 48 | 14.2-18.8 | 16.85 | 0.89 | 20 | 10.7-13.9 | 12.94 | 0.74 | 49 | 12.2-16.6 | 14.51 | 0.88 | ||
| p4 L | 23.9 | 23.9 | 24.3 | 177 | 19-24.6 | 21.97 | 0.97 | 22 | 19-21.9 | 20.68 | 0.77 | 528 | 21-26.8 | 24.19 | 1.01 | 48 | 20.4-26.4 | 24.28 | 1.14 | 23 | 19.7-24.4 | 22.51 | 1.09 | 56 | 21.4-25.2 | 23.14 | 0.99 | ||
| p4 W | 14.9 | 14.9 | 14 | 74 | 10.7-14 | 12.75 | 0.76 | 22 | 11.7-13.6 | 12.73 | 0.50 | 522 | 12.6-17.3 | 15.07 | 0.74 | 49 | 12.7-17.1 | 15.11 | 0.86 | 21 | 9.3-13.8 | 12.88 | 0.92 | 50 | 13.1-16.11 | 14.54 | 0.76 | ||
| m1 L | 24.5 | 24.5 | 176 | 23.3-31.2 | 27.26 | 1.71 | 20 | 23.1-26.5 | 24.91 | 0.91 | 442 | 27.5-38.4 | 32.45 | 1.46 | 47 | 29.7-37.4 | 32.77 | 1.71 | 23 | 22.7-26.3 | 24.33 | 0.99 | 60 | 23-27.6 | 25.26 | 1.26 | |||
| m1 W | 13.0 | 13 | 84 | 9.8-14 | 11.62 | 0.75 | 19 | 10.8-12.3 | 11.52 | 0.48 | 442 | 11.5-15.9 | 13.87 | 0.69 | 46 | 11.7-15.1 | 13.94 | 0.77 | 24 | 10.8-12.82 | 11.83 | 0.56 | 53 | 11-14.81 | 12.82 | 0.81 | |||
| m1tri L | 20.2 | 20.2 | 87 | 21.7-28.2 | 25.20 | 1.53 | 18 | 19.3-22 | 21.03 | 0.79 | 428 | 25.5-33.5 | 29.30 | 1.27 | 35 | 27.2-34.7 | 29.65 | 1.69 | 19 | 18.1-21.2 | 19.64 | 0.90 | 49 | 19-22.8 | 20.91 | 1.02 | |||
| p2 L /m1 L | 0.62 | 0.62 | 171 | 0.47-0.62 | 0.54 | 0.03 | 16 | 0.5-0.63 | 0.56 | 0.03 | 162 | 0.43-0.62 | 0.52 | 0.03 | 29 | 0.43-0.58 | 0.52 | 0.03 | 10 | 0.59-0.76 | 0.65 | 0.05 | 37 | 0.52-0.68 | 0.61 | 0.04 | |||
| p3 W /p3 L | 0.67 | 0.67 | 0.73 | 74 | 0.63-0.79 | 0.71 | 0.03 | 20 | 0.68-0.8 | 0.74 | 0.03 | 505 | 0.6-0.83 | 0.73 | 0.03 | 48 | 0.67-0.79 | 0.73 | 0.03 | 20 | 0.61-0.71 | 0.64 | 0.02 | 49 | 0.58-0.78 | 0.70 | 0.04 | ||
| p3 W /p4 W | 0.97 | 0.97 | 1.19 | 71 | 1.05-1.22 | 1.14 | 0.03 | 19 | 0.99-1.11 | 1.06 | 0.03 | 299 | 0.85-1.2 | 1.10 | 0.04 | 40 | 1.0-1.2 | 1.11 | 0.04 | 20 | 0.95-1.15 | 1.01 | 0.04 | 44 | 0.91-1.11 | 1.01 | 0.03 | ||
| m1tri L /m1 L | 0.82 | 0.82 | 84 | 0.88-0.95 | 0.91 | 0.02 | 17 | 0.81-0.88 | 0.85 | 0.02 | 422 | 0.86-0.95 | 0.90 | 0.02 | 35 | 0.85-0.96 | 0.90 | 0.02 | 18 | 0.76-0.83 | 0.80 | 0.02 | 49 | 0.74-0.87 | 0.83 | 0.02 | |||
| m1tri L /p4 L | 0.85 | 83 | 1.03-1.3 | 1.14 | 0.06 | 18 | 0.9-1.06 | 1.01 | 0.04 | 179 | 1.08-1.37 | 1.21 | 0.05 | 24 | 1.11-1.28 | 1.20 | 0.05 | 13 | 0.79-1.0 | 0.87 | 0.06 | 35 | 0.85-1.02 | 0.90 | 0.04 | ||||
| C L | 16.95 | 16.95 | 40 | 13.4-19 | 16.36 | 1.13 | 6 | 13-15.9 | 14.47 | 0.86 | 31 | 15.1-22.5 | 17.37 | 1.30 | 22 | 16-20.5 | 18.25 | 1.41 | 7 | 17-18.2 | 17.33 | 0.43 | 14 | 16.1-20.8 | 18.61 | 1.25 | |||
| C W | 13.1 | 13.1 | 41 | 10.2-14 | 11.87 | 0.81 | 6 | 10.9-11.9 | 11.35 | 0.33 | 30 | 11.7-15.7 | 13.46 | 0.98 | 22 | 12.5-15.2 | 13.92 | 0.69 | 7 | 12.4-13.1 | 12.66 | 0.24 | 11 | 12.4-15.5 | 13.65 | 0.84 | |||
| P3 L | 22.2 | 22.2 | 190 | 19-25.4 | 21.53 | 1.17 | 7 | 20-21.5 | 20.61 | 0.54 | 374 | 20.7-27.7 | 24.51 | 1.16 | 55 | 19.7-28.3 | 25.38 | 1.62 | 11 | 20.6-23.6 | 22.23 | 0.79 | 55 | 21-25.8 | 23.51 | 1.08 | |||
| P3 W | 16.8 | 16.8 | 80 | 14.2-18.7 | 16.56 | 0.99 | 7 | 14.1-16 | 15.17 | 0.57 | 360 | 15.4-20.5 | 18.21 | 0.91 | 55 | 15.8-21.7 | 19.05 | 1.34 | 8 | 13.73-15.9 | 15.09 | 0.67 | 46 | 14-18.8 | 16.58 | 1.05 | |||
| P4 W | 21.1 | 21.1 | 51 | 16.6-22.5 | 20.28 | 1.36 | 8 | 15-20.4 | 19.08 | 1.65 | 337 | 18-26 | 22.16 | 1.23 | 52 | 19.5-27.9 | 22.57 | 1.78 | 12 | 18.06-22.7 | 19.91 | 1.35 | 47 | 17.9-24.1 | 20.86 | 1.32 | |||
| P4ps L | 11 | 11 | 15 | 6.1-8.9 | 7.48 | 0.74 | 8 | 7.9-9.7 | 8.98 | 0.54 | 29 | 7.4-11.4 | 9.52 | 1.19 | 37 | 7-12.7 | 8.98 | 1.08 | 6 | 9.3-11.3 | 10.43 | 0.71 | 33 | 9.72-13.5 | 11.57 | 1.06 | |||
| P3 W /P3 L | 0.76 | 0.76 | 80 | 0.68-0.86 | 0.75 | 0.03 | 7 | 0.66-0.76 | 0.74 | 0.03 | 345 | 0.67-0.86 | 0.74 | 0.03 | 53 | 0.69-0.83 | 0.75 | 0.04 | 8 | 0.62-0.77 | 0.68 | 0.04 | 46 | 0.6-0.78 | 0.71 | 0.04 | |||
| Huainan, Anhui | Crocuta | Pachycrocuta | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| XLX | DDS | crocuta | honanensis | spelaea | ultima | pyrenaica | perrieri | |||||||||||||||||||
| V15163 | V15160 | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | |
| 1 | 43.8 | 41.2 | 16 | 28-38.5 | 32.36 | 2.74 | 17 | 30.8-39.9 | 36.19 | 3.03 | 32 | 29.1-45 | 38.41 | 4.28 | 21 | 36.6-48.8 | 43.90 | 3.00 | 6 | 36.1-41.4 | 38.93 | 2.11 | 12 | 37.7-44.2 | 41.04 | 1.87 |
| 2 | 48.8 | 16 | 33.8-42.8 | 38.03 | 2.65 | 18 | 31.8-44.8 | 40.30 | 3.56 | 29 | 32.7-52.7 | 43.06 | 5.91 | 21 | 37.9-54.7 | 48.00 | 3.97 | 6 | 41.8-45 | 43.38 | 1.09 | 12 | 38.5-50.8 | 43.88 | 3.69 | |
| 3 | 23.7 | 20.7 | 16 | 15.8-19.3 | 17.37 | 1.00 | 18 | 16.1-20.8 | 18.30 | 1.38 | 36 | 17.8-25.2 | 21.65 | 1.79 | 20 | 19.6-28.2 | 22.77 | 1.90 | 5 | 18.4-21.9 | 19.50 | 1.35 | 13 | 18.1-23.8 | 21.06 | 1.39 |
Table 2 The mandibular measurements and comparisons with other bone-cracking hyenas (mm)
| Huainan, Anhui | Crocuta | Pachycrocuta | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| XLX | DDS | crocuta | honanensis | spelaea | ultima | pyrenaica | perrieri | |||||||||||||||||||
| V15163 | V15160 | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | n | range | X | S | |
| 1 | 43.8 | 41.2 | 16 | 28-38.5 | 32.36 | 2.74 | 17 | 30.8-39.9 | 36.19 | 3.03 | 32 | 29.1-45 | 38.41 | 4.28 | 21 | 36.6-48.8 | 43.90 | 3.00 | 6 | 36.1-41.4 | 38.93 | 2.11 | 12 | 37.7-44.2 | 41.04 | 1.87 |
| 2 | 48.8 | 16 | 33.8-42.8 | 38.03 | 2.65 | 18 | 31.8-44.8 | 40.30 | 3.56 | 29 | 32.7-52.7 | 43.06 | 5.91 | 21 | 37.9-54.7 | 48.00 | 3.97 | 6 | 41.8-45 | 43.38 | 1.09 | 12 | 38.5-50.8 | 43.88 | 3.69 | |
| 3 | 23.7 | 20.7 | 16 | 15.8-19.3 | 17.37 | 1.00 | 18 | 16.1-20.8 | 18.30 | 1.38 | 36 | 17.8-25.2 | 21.65 | 1.79 | 20 | 19.6-28.2 | 22.77 | 1.90 | 5 | 18.4-21.9 | 19.50 | 1.35 | 13 | 18.1-23.8 | 21.06 | 1.39 |
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 | PC 8 | PC 9 | PC 10 | PC 11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| c L | 0.12857 | 0.52793 | -0.51798 | -0.4058 | -0.1319 | 0.42808 | -0.10251 | 0.18228 | -0.14816 | 0.0031197 | 0.073732 |
| c W | 0.11519 | 0.41137 | -0.2895 | 0.061824 | 0.19053 | -0.40619 | 0.5479 | -0.3734 | 0.27855 | 0.028902 | 0.10265 |
| p2 L | 0.19562 | 0.21439 | 0.60374 | -0.35355 | 0.019271 | 0.090964 | 0.19582 | -0.23824 | -0.34804 | 0.44345 | 0.068881 |
| p2 W | 0.20677 | 0.14561 | 0.35952 | -0.015387 | 0.3512 | 0.43567 | 0.23062 | 0.25402 | 0.47061 | -0.33123 | -0.21097 |
| p3 L | 0.26228 | 0.23653 | 0.21418 | -0.34127 | -0.051144 | -0.48927 | -0.49421 | -0.095557 | 0.074532 | -0.45689 | -0.057054 |
| p3 W | 0.22325 | 0.10562 | -0.01265 | 0.26965 | 0.33499 | -0.22746 | 0.19058 | 0.45676 | -0.63522 | -0.22854 | 0.038555 |
| p4 L | 0.18885 | 0.25885 | 0.20094 | 0.31979 | -0.72407 | -0.12879 | 0.13943 | 0.37982 | 0.18512 | 0.10872 | 0.063554 |
| p4 W | 0.1571 | 0.28103 | -0.066492 | 0.34547 | 0.27647 | -0.061683 | -0.42603 | 0.025896 | 0.09434 | 0.51327 | -0.48766 |
| m1 L | 0.56868 | -0.22759 | -0.14619 | 0.18498 | -0.26791 | 0.2328 | 0.1042 | -0.44614 | -0.21226 | -0.22508 | -0.37099 |
| m1 W | 0.16425 | 0.16438 | 0.10107 | 0.44869 | 0.12938 | 0.26572 | -0.32376 | -0.2717 | 0.024521 | -0.081191 | 0.67917 |
| m1tri L | 0.59986 | -0.43932 | -0.17806 | -0.24483 | 0.13613 | -0.13176 | -0.023896 | 0.26553 | 0.24489 | 0.31513 | 0.29878 |
| Eigenvalue | 29.9412 | 5.66701 | 0.970645 | 0.672578 | 0.502124 | 0.342199 | 0.295265 | 0.200275 | 0.181543 | 0.150677 | 0.108996 |
| % variance | 76.708 | 14.519 | 2.4868 | 1.7231 | 1.2864 | 0.8767 | 0.75646 | 0.5131 | 0.46511 | 0.38603 | 0.27924 |
Table 3 Principal component loadings and percentage of variance explained for PCA including the canine
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 | PC 8 | PC 9 | PC 10 | PC 11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| c L | 0.12857 | 0.52793 | -0.51798 | -0.4058 | -0.1319 | 0.42808 | -0.10251 | 0.18228 | -0.14816 | 0.0031197 | 0.073732 |
| c W | 0.11519 | 0.41137 | -0.2895 | 0.061824 | 0.19053 | -0.40619 | 0.5479 | -0.3734 | 0.27855 | 0.028902 | 0.10265 |
| p2 L | 0.19562 | 0.21439 | 0.60374 | -0.35355 | 0.019271 | 0.090964 | 0.19582 | -0.23824 | -0.34804 | 0.44345 | 0.068881 |
| p2 W | 0.20677 | 0.14561 | 0.35952 | -0.015387 | 0.3512 | 0.43567 | 0.23062 | 0.25402 | 0.47061 | -0.33123 | -0.21097 |
| p3 L | 0.26228 | 0.23653 | 0.21418 | -0.34127 | -0.051144 | -0.48927 | -0.49421 | -0.095557 | 0.074532 | -0.45689 | -0.057054 |
| p3 W | 0.22325 | 0.10562 | -0.01265 | 0.26965 | 0.33499 | -0.22746 | 0.19058 | 0.45676 | -0.63522 | -0.22854 | 0.038555 |
| p4 L | 0.18885 | 0.25885 | 0.20094 | 0.31979 | -0.72407 | -0.12879 | 0.13943 | 0.37982 | 0.18512 | 0.10872 | 0.063554 |
| p4 W | 0.1571 | 0.28103 | -0.066492 | 0.34547 | 0.27647 | -0.061683 | -0.42603 | 0.025896 | 0.09434 | 0.51327 | -0.48766 |
| m1 L | 0.56868 | -0.22759 | -0.14619 | 0.18498 | -0.26791 | 0.2328 | 0.1042 | -0.44614 | -0.21226 | -0.22508 | -0.37099 |
| m1 W | 0.16425 | 0.16438 | 0.10107 | 0.44869 | 0.12938 | 0.26572 | -0.32376 | -0.2717 | 0.024521 | -0.081191 | 0.67917 |
| m1tri L | 0.59986 | -0.43932 | -0.17806 | -0.24483 | 0.13613 | -0.13176 | -0.023896 | 0.26553 | 0.24489 | 0.31513 | 0.29878 |
| Eigenvalue | 29.9412 | 5.66701 | 0.970645 | 0.672578 | 0.502124 | 0.342199 | 0.295265 | 0.200275 | 0.181543 | 0.150677 | 0.108996 |
| % variance | 76.708 | 14.519 | 2.4868 | 1.7231 | 1.2864 | 0.8767 | 0.75646 | 0.5131 | 0.46511 | 0.38603 | 0.27924 |
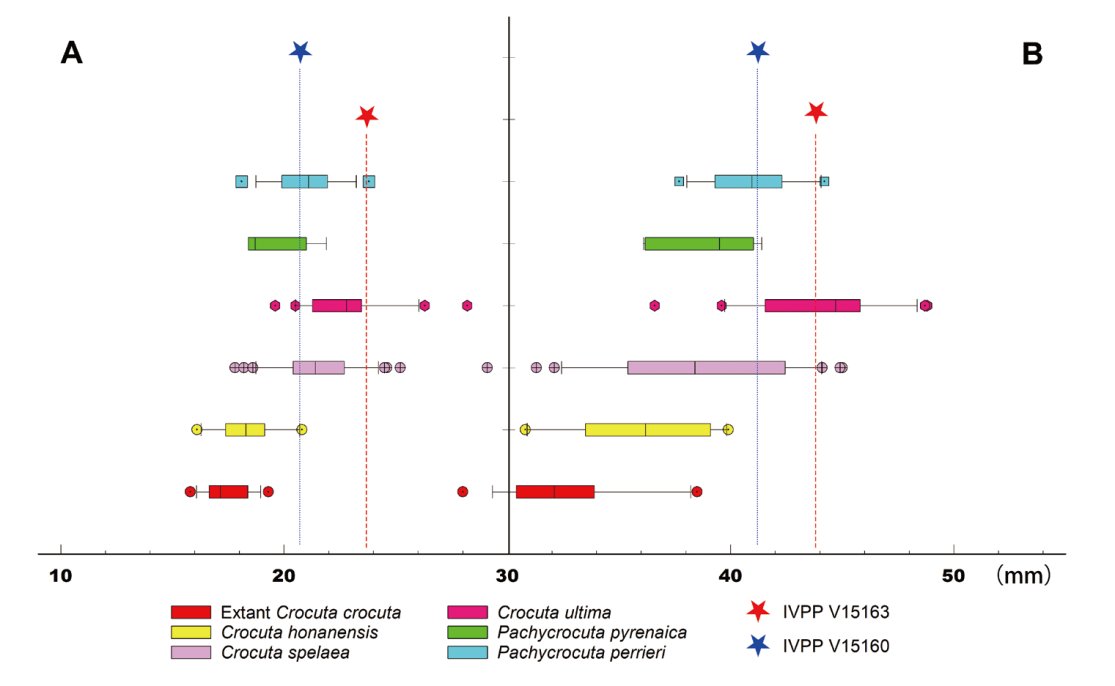
Fig. 3 Box plots of mandibular thickness and depth in hyenas A. the thickness measured below p3; B. the depth measured anterior to p3 The data used for plotting were taken from Table 2
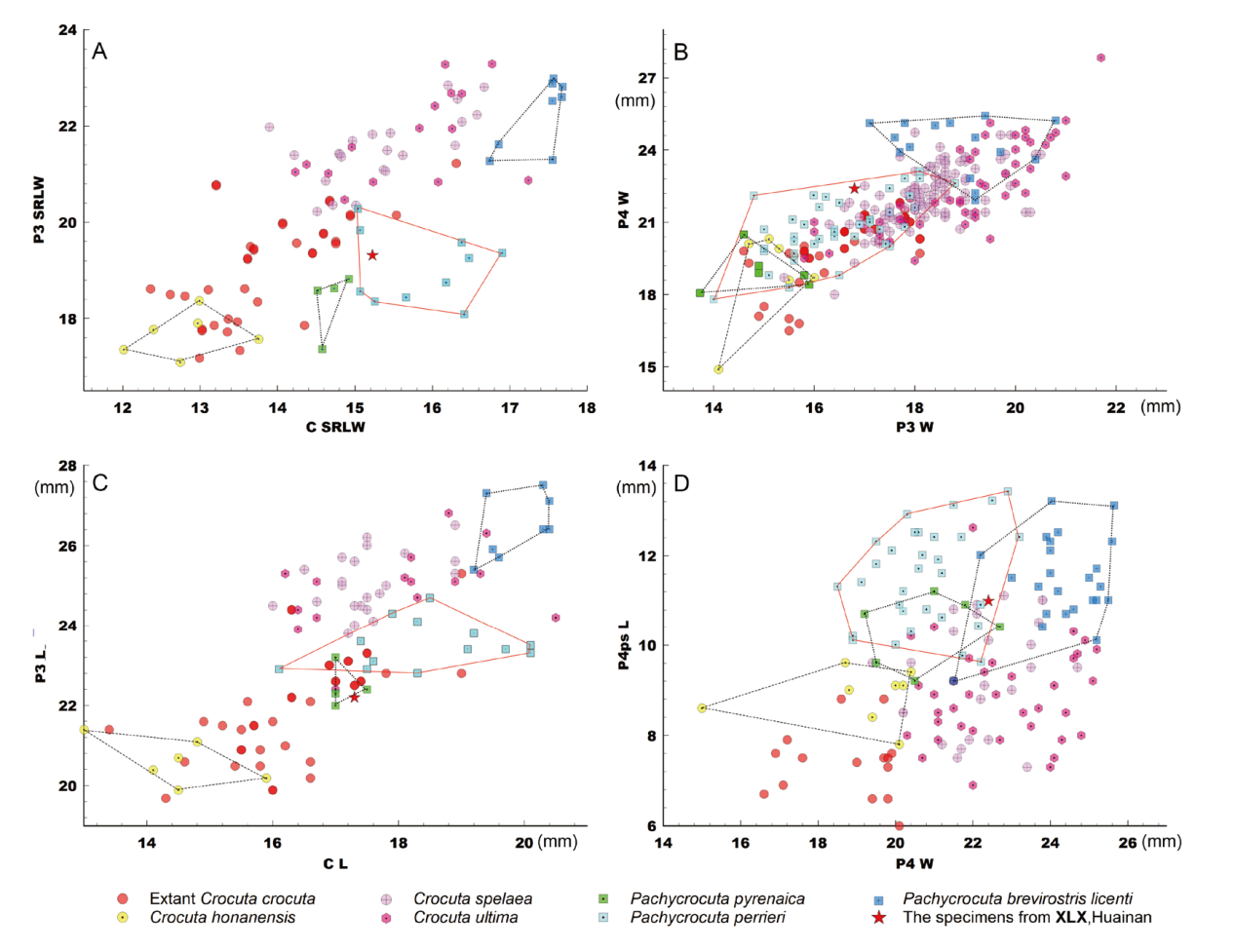
Fig. 4 Bivariate plots of the upper teeth in hyenas The data used to create the plots or their sources are detailed in Supplementary Tables 2, 3
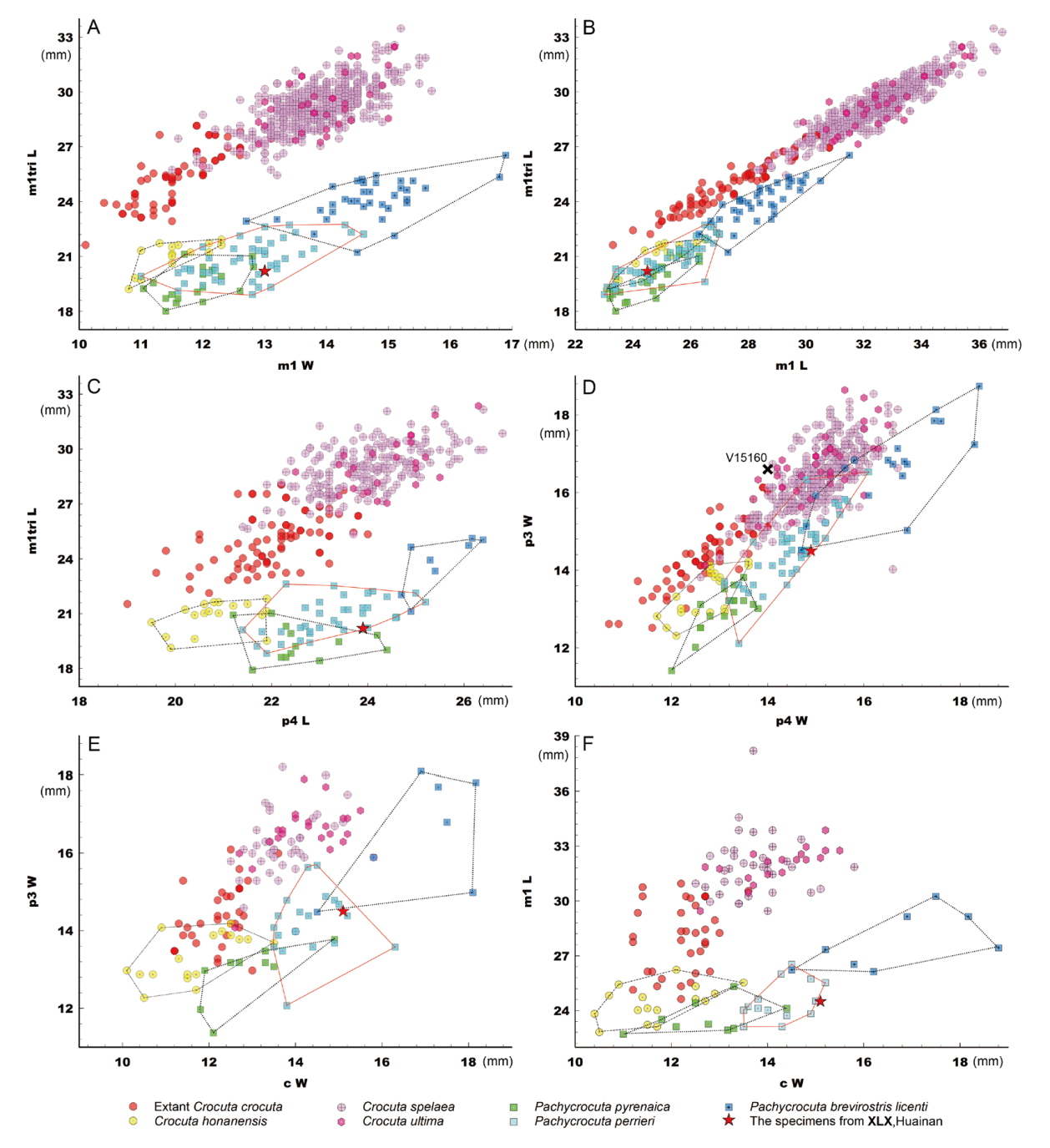
Fig. 5 Bivariate plots of the lower teeth in hyenas The data used to create the plots or their sources are detailed in Supplementary Tables 1, 3 Xiliexi (XLX) specimens are sporadically fragmented, without a complete lower dentition attached, so much so that IVPP V15163 and V15164.3-6 had to be combined and virtualized as a single sample here (calculated by averaging)
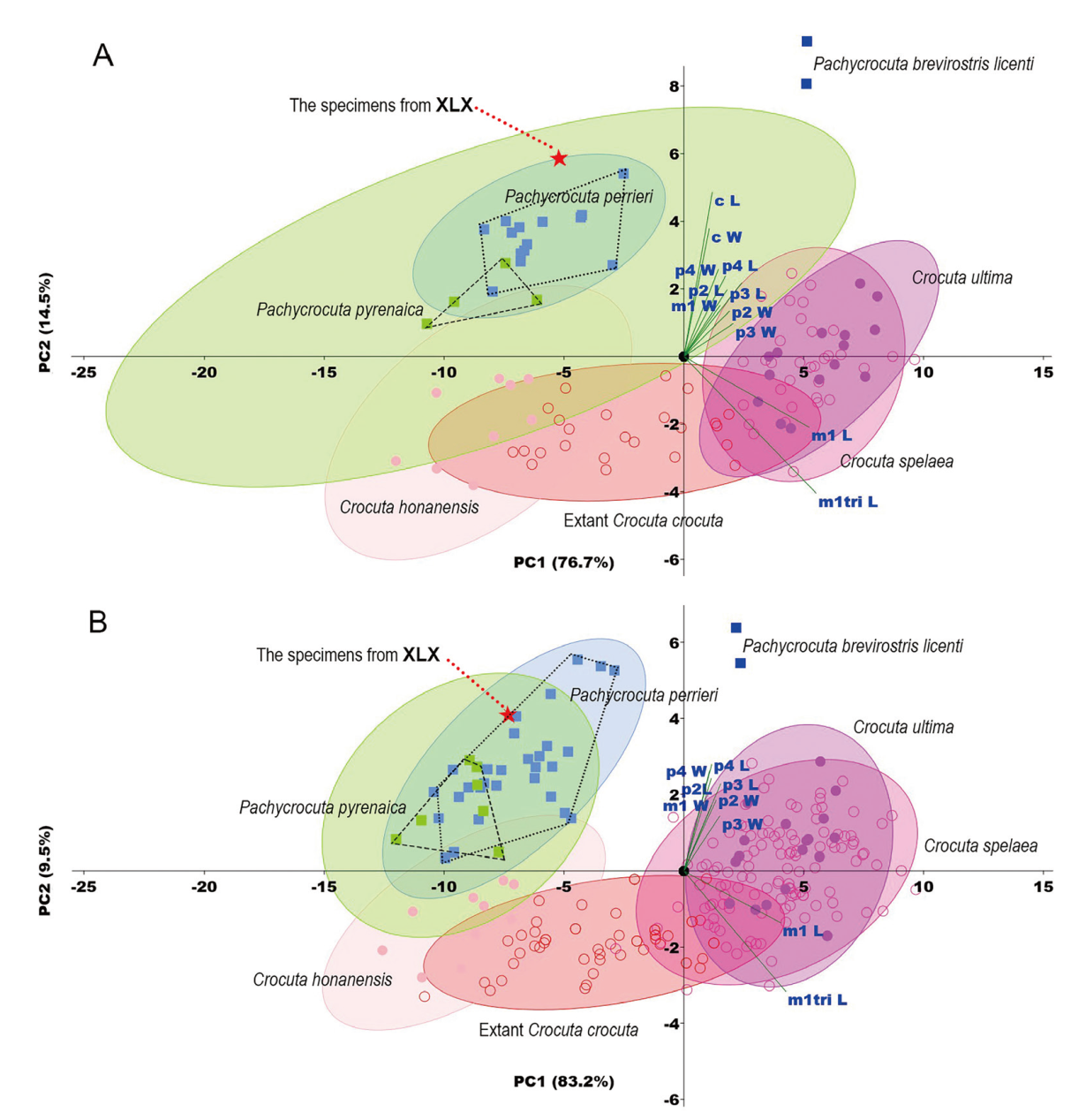
Fig. 6 The Principal Component Analysis (PCA) plot of hyenas based on selected lower tooth variables A. PCA including the lower canine; B. PCA without the lower canine The data used to create the plots or their sources are detailed in Supplementary Tables 1, 3 The ellipses in the figure have a 95% confidence level Details of the parameters of the PCA are given in Tables 3, 4 Xiliexi (XLX) specimens are sporadically fragmented, without a complete lower dentition attached, so much so that IVPP V15163 and V15164.3-6 had to be combined and virtualized as a single sample (calculated by averaging)
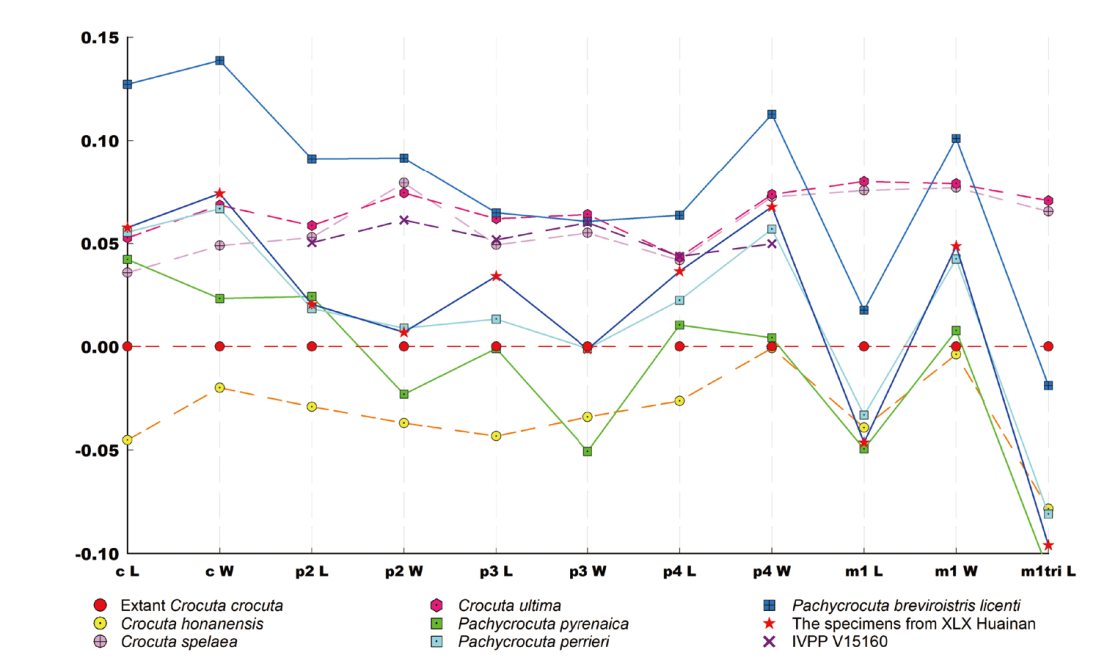
Fig. 7 Log-ratio plots of tooth measurements (length and width) in hyenas The extant Crocuta crocuta is taken as the reference baseline, the data used to create the plots or their sources are detailed in Supplementary Tables 1, 3 All categories shown in the figure were calculated by averaging the individuals they contain, except for V15160, the only individual in the plot
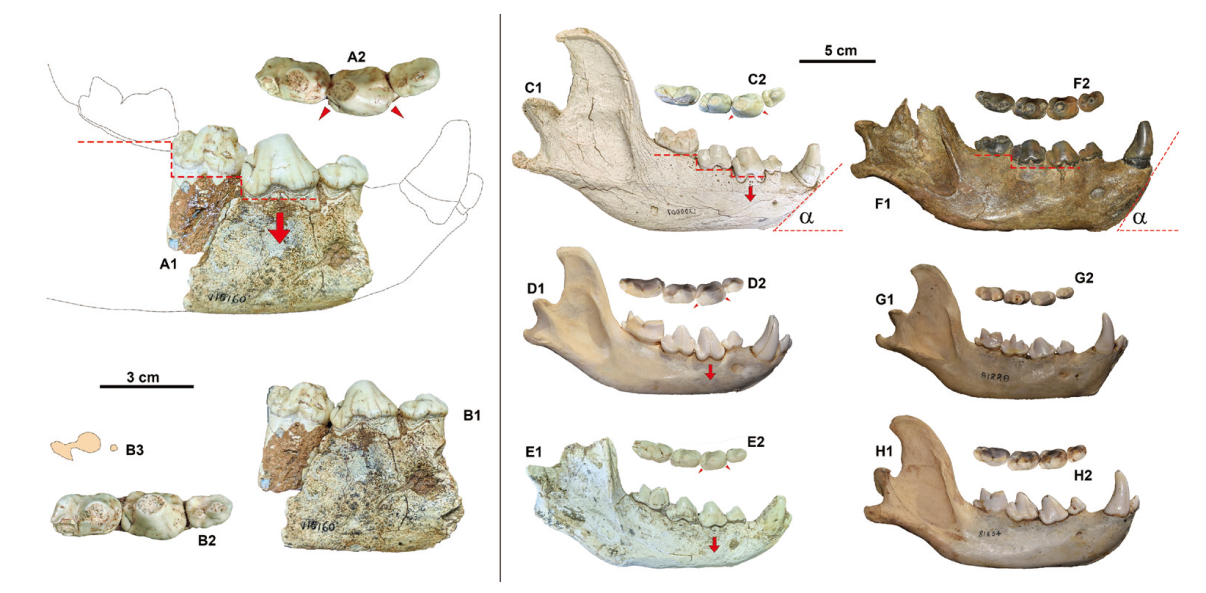
Fig. 8 Morphological comparisons of hyena mandibles A, B. Crocuta cf. C. ultima (IVPP V15160) from DDS of Huainan: A1. a virtual mandibular outline based on the reassembled specimens in buccal view, A2. the reassembled dentition in occlusal view; B. an original incorrectly joined specimen: B1. buccal view, B2. occlusal view, B3. an outline of the abrasive surface of p4; C. Crocuta ultima (LXD0007); D. C. crocuta (AMNH M187773); E. C. honanensis (IVPP V32018); F. Pachycrocuta perrieri (F:AM107767); G. Hyaena hyaena (AMNH M81228); H. Parahyaena brunnea (AMNH M81834); 1. buccal views; 2. occlusal views The short-stalked red arrows indicate that p3 of Crocuta significantly lower than any other check teeth, so much so that p3, p4 and m1 look like being placed on an ascending staircase (see the red dashed line under the teeth of A1 and C1, D1, E1?). The small triangles show that p3 of Crocuta is so dilated, especially on the buccal side, as that its buccal margin protrudes clearly buccally over the other cheek teeth (see A2, C2, D2 and E2). The angles (α) in C1 and F1 are 44.20º and 54.40º, respectively
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 | PC 8 | PC 9 | |
|---|---|---|---|---|---|---|---|---|---|
| p2 L | 0.16873 | 0.35737 | -0.71404 | 0.20207 | -0.12748 | -0.15906 | 0.49526 | 0.041134 | 0.067926 |
| p2 W | 0.1992 | 0.27239 | -0.19687 | 0.52563 | -0.091612 | 0.14415 | -0.71089 | -0.042939 | -0.18629 |
| p3 L | 0.22679 | 0.33633 | -0.11715 | -0.3716 | 0.76766 | -0.1924 | -0.20981 | -0.060952 | 0.0979 |
| p3 W | 0.22118 | 0.21119 | 0.2886 | 0.28189 | 0.20924 | 0.63822 | 0.32283 | -0.3378 | 0.27257 |
| p4 L | 0.17126 | 0.41049 | -0.012391 | -0.6216 | -0.45592 | 0.39385 | -0.12733 | 0.19038 | 0.013513 |
| p4 W | 0.15844 | 0.38652 | 0.44594 | 0.12696 | 0.042212 | -0.19795 | 0.27951 | 0.21256 | -0.66802 |
| m1 L | 0.59493 | -0.20026 | 0.061079 | -0.1577 | -0.27039 | -0.30134 | -0.0061062 | -0.63567 | -0.096583 |
| m1 W | 0.17113 | 0.25303 | 0.38188 | 0.18844 | -0.20168 | -0.43475 | -0.069673 | 0.26436 | 0.64879 |
| m1tri L | 0.62757 | -0.46727 | -0.076152 | 0.049466 | 0.13727 | 0.18832 | 0.0435 | 0.56855 | -0.01097 |
| Eigenvalue | 28.0402 | 3.21113 | 0.742061 | 0.569183 | 0.441519 | 0.25669 | 0.17803 | 0.143479 | 0.134145 |
| % variance | 83.165 | 9.5239 | 2.2009 | 1.6881 | 1.3095 | 0.76132 | 0.52802 | 0.42555 | 0.39786 |
Table 4 Principal component loadings and percentage of variance explained for PCA excluding the canine
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 | PC 8 | PC 9 | |
|---|---|---|---|---|---|---|---|---|---|
| p2 L | 0.16873 | 0.35737 | -0.71404 | 0.20207 | -0.12748 | -0.15906 | 0.49526 | 0.041134 | 0.067926 |
| p2 W | 0.1992 | 0.27239 | -0.19687 | 0.52563 | -0.091612 | 0.14415 | -0.71089 | -0.042939 | -0.18629 |
| p3 L | 0.22679 | 0.33633 | -0.11715 | -0.3716 | 0.76766 | -0.1924 | -0.20981 | -0.060952 | 0.0979 |
| p3 W | 0.22118 | 0.21119 | 0.2886 | 0.28189 | 0.20924 | 0.63822 | 0.32283 | -0.3378 | 0.27257 |
| p4 L | 0.17126 | 0.41049 | -0.012391 | -0.6216 | -0.45592 | 0.39385 | -0.12733 | 0.19038 | 0.013513 |
| p4 W | 0.15844 | 0.38652 | 0.44594 | 0.12696 | 0.042212 | -0.19795 | 0.27951 | 0.21256 | -0.66802 |
| m1 L | 0.59493 | -0.20026 | 0.061079 | -0.1577 | -0.27039 | -0.30134 | -0.0061062 | -0.63567 | -0.096583 |
| m1 W | 0.17113 | 0.25303 | 0.38188 | 0.18844 | -0.20168 | -0.43475 | -0.069673 | 0.26436 | 0.64879 |
| m1tri L | 0.62757 | -0.46727 | -0.076152 | 0.049466 | 0.13727 | 0.18832 | 0.0435 | 0.56855 | -0.01097 |
| Eigenvalue | 28.0402 | 3.21113 | 0.742061 | 0.569183 | 0.441519 | 0.25669 | 0.17803 | 0.143479 | 0.134145 |
| % variance | 83.165 | 9.5239 | 2.2009 | 1.6881 | 1.3095 | 0.76132 | 0.52802 | 0.42555 | 0.39786 |
| Pachycrocuta | Crocuta | ||
|---|---|---|---|
| PF and APF | I-1 | PF ovoid or rounded rectangular, with a symmetrical and almost parallel configuration | PF comparatively narrow, lanceolate in shape, with distinctly divergent distal ends |
| I-2 | APF relatively long, more than half the total length of PF | APF slightly short, less than half the total length of PF | |
| incisor | II-1 | I3 notably large, almost twice the size of I1 | I3 relatively weak, rarely more than twice the size of I1 |
| upper/lower canine | III-1 | large with prominent anterior and posterior lateral crests | relatively small with inconspicuous anterior and posterior lateral crests |
| III-2 | cingulum so developed around the base that the crown and roots are clearly separated | cingulum less developed, the crown and roots not clearly separated | |
| III-3 | a triangular protuberance better developed at the intersection of the cingulum and the anterolateral crest on the upper canine | no protuberances developed at all | |
| III-4 | almost upright implanted in maxilla or mandible | somewhat backward tilted implantation of the maxilla or mandible | |
| P2/p2 | IV-1 | relatively large, P2L/P4L and p2L/m1L normally larger than 0.5 and 0.6 respectively | much reduced, P2L/P4L and p2L/m1L barely larger than 0.5 and 0.6 respectively |
| IV-2 | a rounded rectangle with the anterior and posterior ends nearly equal in width | a quadrilateral with the posterior end normally wider than the anterior | |
| IV-3 | the main cusp is high and large with a small PAC | in contrast, the main cusp is slightly low and small with a large PAC | |
| P3/p3 | V-1 | crown not particularly high and wide, its ratio of W/L somewhat lower | crown especially high and wide, typical hypsodont |
| V-2 | AAC slightly or barely developed, PAC normally developed as those of other premolars | AAC barely developed, PAC often high and short anterior-posteriorly | |
| V-3 | two-element structure, namely both PAC and the posterior cingulum/cingulid present | single-element structure, namely either PAC or cingulum/cingulid present | |
| V-4 | p3 comparable to p4 in width, or even narrower than the latter | p3 nearly always larger than p4 in width | |
| P4 | VI-1 | relatively short but massive, especially for the metastyle | long and slim, with an extremely long metastyle |
| VI-2 | the protocone large with a normal position | the protocone large or small, significantly tilted forward beyond the front edge of the parastyle | |
| VI-3 | the space lingually located between P3 and P4 is normally broad | the space quite narrow, almost non-existent | |
| VI-4 | parastyle relatively large with a conical shape, almost comparable to paracone | parastyle distinctly smaller than paracone with a tongue-shaped form | |
| p4 | VII-1 | main cusp with both AAC and PAC developed, AAC slightly smaller than PAC, but always present | AAC highly reduced and even totally disappeared in some species |
| VII-2 | PAC centrally located, while the terrace on the lingual side quite narrow | PAC strongly labially biased, forming a wide terrace or basin on the lingual side | |
| M1 | VIII-1 | always present and relatively large with three distinct roots | either absent or present as small one normally with 1-2 roots |
| m1 | IX-1 | quite short and massive, especially the paraconid that is distinctly wider than the protoconid | long and slender, almost crescent-shaped, the paraconid almost comparable to the protoconid in width |
| IX-2 | the anterior edge of the paraconid clearly tilted posteriorly | the anterior edge of the paraconid nearly erect, nearly parallel to posterior edge of the protoconid | |
| IX-3 | the metaconid reduced, but often present in the primitive species | the metaconid highly reduced, almost completely disappeared | |
| IX-4 | the talonid relatively large, often larger than 15% of the total length | the talonid small, barely larger than 15% of the total length | |
| IX-5 | the talonid of complicated structure, normally with 2 or 3 cuspids on it | the talonid quite simple with 1 or 2 cuspids on it in most cases | |
| mandible | X-1 | ramus relatively massive, both deep and thick | ramus less in both depth and thickness |
| X-2 | the dorsal and ventral margins almost parallel with a comparable depth, except slightly increase at canine and m1 | The dorsal margin exhibiting an upward trajectory from the proximal to the distal end, accompanied by a substantial deepening at m1 | |
| X-3 | the anterior plane of the symphysis slightly upright, forming a larger angle with the ventral margin | the anterior plane clearly tilted backward, forming a smaller angle | |
| X-4 | all cheek teeth, except for the carnassial, nearly on the same plane (labial view) | p3 significantly lower than other teeth, as if set at the bottom step of a staircase | |
| X-5 | the dentition straight or slightly curved in an arc shape (crown view) | the dentition forming a strongly curved arc with p3 clearly protruding beyond others |
Table 5 Differences in dentognathic morphology between Pachycrocuta and Crocuta
| Pachycrocuta | Crocuta | ||
|---|---|---|---|
| PF and APF | I-1 | PF ovoid or rounded rectangular, with a symmetrical and almost parallel configuration | PF comparatively narrow, lanceolate in shape, with distinctly divergent distal ends |
| I-2 | APF relatively long, more than half the total length of PF | APF slightly short, less than half the total length of PF | |
| incisor | II-1 | I3 notably large, almost twice the size of I1 | I3 relatively weak, rarely more than twice the size of I1 |
| upper/lower canine | III-1 | large with prominent anterior and posterior lateral crests | relatively small with inconspicuous anterior and posterior lateral crests |
| III-2 | cingulum so developed around the base that the crown and roots are clearly separated | cingulum less developed, the crown and roots not clearly separated | |
| III-3 | a triangular protuberance better developed at the intersection of the cingulum and the anterolateral crest on the upper canine | no protuberances developed at all | |
| III-4 | almost upright implanted in maxilla or mandible | somewhat backward tilted implantation of the maxilla or mandible | |
| P2/p2 | IV-1 | relatively large, P2L/P4L and p2L/m1L normally larger than 0.5 and 0.6 respectively | much reduced, P2L/P4L and p2L/m1L barely larger than 0.5 and 0.6 respectively |
| IV-2 | a rounded rectangle with the anterior and posterior ends nearly equal in width | a quadrilateral with the posterior end normally wider than the anterior | |
| IV-3 | the main cusp is high and large with a small PAC | in contrast, the main cusp is slightly low and small with a large PAC | |
| P3/p3 | V-1 | crown not particularly high and wide, its ratio of W/L somewhat lower | crown especially high and wide, typical hypsodont |
| V-2 | AAC slightly or barely developed, PAC normally developed as those of other premolars | AAC barely developed, PAC often high and short anterior-posteriorly | |
| V-3 | two-element structure, namely both PAC and the posterior cingulum/cingulid present | single-element structure, namely either PAC or cingulum/cingulid present | |
| V-4 | p3 comparable to p4 in width, or even narrower than the latter | p3 nearly always larger than p4 in width | |
| P4 | VI-1 | relatively short but massive, especially for the metastyle | long and slim, with an extremely long metastyle |
| VI-2 | the protocone large with a normal position | the protocone large or small, significantly tilted forward beyond the front edge of the parastyle | |
| VI-3 | the space lingually located between P3 and P4 is normally broad | the space quite narrow, almost non-existent | |
| VI-4 | parastyle relatively large with a conical shape, almost comparable to paracone | parastyle distinctly smaller than paracone with a tongue-shaped form | |
| p4 | VII-1 | main cusp with both AAC and PAC developed, AAC slightly smaller than PAC, but always present | AAC highly reduced and even totally disappeared in some species |
| VII-2 | PAC centrally located, while the terrace on the lingual side quite narrow | PAC strongly labially biased, forming a wide terrace or basin on the lingual side | |
| M1 | VIII-1 | always present and relatively large with three distinct roots | either absent or present as small one normally with 1-2 roots |
| m1 | IX-1 | quite short and massive, especially the paraconid that is distinctly wider than the protoconid | long and slender, almost crescent-shaped, the paraconid almost comparable to the protoconid in width |
| IX-2 | the anterior edge of the paraconid clearly tilted posteriorly | the anterior edge of the paraconid nearly erect, nearly parallel to posterior edge of the protoconid | |
| IX-3 | the metaconid reduced, but often present in the primitive species | the metaconid highly reduced, almost completely disappeared | |
| IX-4 | the talonid relatively large, often larger than 15% of the total length | the talonid small, barely larger than 15% of the total length | |
| IX-5 | the talonid of complicated structure, normally with 2 or 3 cuspids on it | the talonid quite simple with 1 or 2 cuspids on it in most cases | |
| mandible | X-1 | ramus relatively massive, both deep and thick | ramus less in both depth and thickness |
| X-2 | the dorsal and ventral margins almost parallel with a comparable depth, except slightly increase at canine and m1 | The dorsal margin exhibiting an upward trajectory from the proximal to the distal end, accompanied by a substantial deepening at m1 | |
| X-3 | the anterior plane of the symphysis slightly upright, forming a larger angle with the ventral margin | the anterior plane clearly tilted backward, forming a smaller angle | |
| X-4 | all cheek teeth, except for the carnassial, nearly on the same plane (labial view) | p3 significantly lower than other teeth, as if set at the bottom step of a staircase | |
| X-5 | the dentition straight or slightly curved in an arc shape (crown view) | the dentition forming a strongly curved arc with p3 clearly protruding beyond others |
| [1] | Baryshnikov G, 2014. Late Pleistocene hyena Crocuta ultima ussurica (Mammalia, Carnivora, Hyaenidae) from the Paleolithic site in Geographical Society Cave in the Russian Far East. Proc Zool Inst RAS, 318(3): 197-225 |
| [2] | Baryshnikov G, Tsoukala E, 2010. New analysis of the Pleistocene carnivores from Petralona cave (Macedonia, Greece) based on the collection of the Thessaloniki Aristotle university. Geobios, 43: 389-402 |
| [3] | Boaz N T, Ciochon R L, Xu Q Q et al., 2000. Large mammalian carnivores as a taphonomic factor in the bone accumulation at Zhoukoudian. Acta Anthrop Sin, 19: 224-234 |
| [4] | Boaz N T, Ciochon R L, Xu Q Q et al., 2004. Mapping and taphonomic analysis of the Homo erectus loci at Locality 1 Zhoukoudian, China. J Hum Evol, 46(5): 519-549 |
| [5] | Bonifay M F, 1971. Carnivores Quaternaires du Sud-Est de la France. Mém Mus Natl Hist Nat, C(21): 1-358 |
| [6] | Boule M, 1893. Description de l’Hyaena brevirostris du Pliocene de Sainzelles pres le Puy (Haute-Loire). Ann Sci Nat, 8(15): 85-97 |
| [7] | Busk G, 1868. Remarks on the cranial and dental characters of the existing species of Hyaena. J Linn Soc, 9(59): 59-79 |
| [8] | Chen S K, Pei J, Yi J et al., 2017. Preliminary report on the mammalian fauna from Yumidong Cave, Wushan, Chongqing, and its chronological analysis. Quat Sci, 37(4): 845-852 |
| [9] | Chen Z W, Li J J, Yu S F, 2001. A Paleolithic site at Chuanfandong in Sanming City, Fujian Province. Acta Anthrop Sin, 20: 256-270 |
| [10] | Chia L P, 1957. Notes on the human and some other mammalian remains from Changyang, Hupei. Vert PalAsiat, 1: 247-257 |
| [11] | Colbert E H, Hooijer D A, 1953. Pleistocene mammals from the limestone fissures of Szechwan, China. Bull Am Mus Nat Hist, 102(1): 1-134 |
| [12] | Dennell E W, Coard R, Turner A, 2008. Predators and scavengers in Early Pleistocene southern Asia. Quat Int, 192: 78-88 |
| [13] | Dong W, 2006. Early Pleistocene ruminants (mammals) from the Dajushan, Huainan, Anhui Province (China). Vert PalAsiat, 44: 332-346 |
| [14] | Dong W, 2008. Early Pleistocene suid (mammal) from the Dajushan, Huainan, Anhui Province (China). Vert PalAsiat, 46: 233-246 |
| [15] | Ewer R F, 1954a. Some adaptive features in the dentition of hyaenas. Ann Mag Nat Hist, 7: 188-194 |
| [16] | Ewer R F, 1954b. The fossil carnivores of the Transvaal caves. The Hyaenidae of Kromdraai. Proc Zool Soc London, 124: 565-585 |
| [17] | Ewer R F, 1973. The Carnivores. Ithaca, New York: Cornell University Press. 1-494 |
| [18] | Fang Y S, 2000. Mammalian fauna since the Pliocene in Jiangsu. Southeast Culture, 135: 33-40 |
| [19] | Galobart A, Antón A, Maroto J, 2003. Hiénidos y cánidos de los yacimientos de Incarcal (Girona, NE de la Península Ibérica). Una aproximación a la paleobiología del hiénido del Pleistoceno inferior. Paleontol Evol, 34: 79-98 |
| [20] | Hammer Ø, Harper D, Ryan P, 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron, 4(1): 1-9 |
| [21] | Han D F, Xu C H, 1989. Quaternary mammalian faunas and environment of fossil humans in South China. In: Wu R K, Wu X Z, Zhang S S eds. Early Humankind in China. Beijing: Science Press. 338-391 |
| [22] | Han D F, Zhang S S, 1978. A hominid canine and mammalian material from the Pleistocene of Zhejiang. Vert PalAsiat, 16: 255-263 |
| [23] | Holekamp K E, Kolowski J M, 2009. Family Hyaenidae (hyaenas). In: Wilson D E, Mittermeier R A eds. Handbook of the Mammals of the World. Barcelona: Lynx Ediciones. 234-262 |
| [24] | Holekamp K E, Smale L, 1990. Provisioning and food sharing by lactating spotted hyenas (Crocuta crocuta). Ethology, 86: 191-202 |
| [25] | Howell F C, Petter G, 1980. The Pachycrocuta and Hyaena lineages (Plio-Pleistocene and extant species of the Hyaenidae). Their relationships with Miocene Ictitheres: Palhyaena and Hyaenictitherium. Geobios, 13: 579-623 |
| [26] | Hu C K, 1985. The history of mammalian fauna of Locality 1 of Zhoukoudian and its recent advances. In: Wu R K, Ren M, Zhu X M et al. eds. Multi-disciplinary Study of the Peking Man Site at Zhoukoudian. Beijing: Science Press. 107-113 |
| [27] | Huang W P, 1989. Taxonomy of the Hyaenidae (Hyaena and Crocuta) of the Pleistocene in China. Verte PalAsiat, 27: 197-204 |
| [28] | Huang W P, Fang D S, Ye Y X, 1982. Preliminary study of the hominid skull and fauna of Hexian, Anhui. Vert PalAsiat, 20: 248-256 |
| [29] | Iannucci A, Mecozzi B, Sardella R et al., 2021. The extinction of the giant hyena Pachycrocuta brevirostris and a reappraisal of the Epivillafranchian and Galerian Hyaenidae in Europe: faunal turnover during the Early-Middle Pleistocene Transition. Quat Sci Rev, 272: 107240 |
| [30] | Jiangzuo Q G, Liu J Y, Jiang H et al., 2022. A huge Pachycrocuta from the middle Pleistocene loess in Luoning county, central China, and the evolution of mandible within Pliocrocuta-Pachycrocuta lineage. Hist Biol, doi: 10.1080/08912963.2022.2056839 |
| [31] | Jin C Z, 2004. Fossil leporids (Mammalia, Lagomorpha) from Huainan, Anhui, China. Vert PalAsiat, 42: 230-245 |
| [32] | Jin C Z, Yoshinari K, Hiroyuk T, 1999. Pliocene and Early Pleistocene insectivore and rodent faunas from Dajushan, Qipanshan and Haimao in North China and the reconstruction of the faunal succession from the Late Miocene to Middle Pleistocene. J Geosci, 42(1): 1-19 |
| [33] | Kowalski K, 1976. Mammals: An Outline of Theriology. Warszawa: Polish Scientific Publishers. 1-617 |
| [34] | Kretzoi M, 1938. Die Raubtiere von Gombaszög nebst einer Übersicht der Gesamtfauna. Ann Mus Nat Hungarici, 31: 89-157 |
| [35] | Kruuk H, 1972. The Spotted Hyaena. Chicago: University of Chicago Press. 1-335 |
| [36] | Kurtén B, 1956. The status and affinities of Hyaena sinensis Owen and Hyaena ultima Matsumoto. Am Mus Novit, 1764: 1-48 |
| [37] | Kurtén B, 1957. The bears and hyaenas of the interglacials. Quaternaria, 4: 1-13 |
| [38] | Kurtén B, 1958. A note on the hyaenid remains from the Lungtung Cave described by Chia. Vert PalAsiat, 2: 164 |
| [39] | Kurtén B, 1968. Pleistocene Mammals of Europe. London: Weidenfeld and Nicolson. 1-317 |
| [40] | Kurtén B, 1972. Fossil Hyaenidae from the excavations at Stránská Skála. Anthropos, 20: 113-120 |
| [41] | Kurtén B, Garevski R, 1989. Giant hyena, Hyaena brevirostris Aymard (Mammalia, Carnivora), from the Middle Pleistocene of Manastirec, Yugoslavia. Ann Zool Fenn, 26(2): 45-147 |
| [42] | Lewis M, Werdelin L, 2022. A revision of the genus Crocuta (Mammalia, Hyaenidae). Palaeontogr Abt A, 322: 1-115 |
| [43] | Li W M, Zhang Z F, Gu Y M et al., 1982. A fauna from Lianhua cave, Dantu, Jiangsu. Acta Anthrop Sin, 1: 169-214 |
| [44] | Li Y, Liu W H, Wang L H, 2021. Chinese Machairodontinae (Carnivora, Felidae) and their ecological significance. Chinese Sci Bull, 66: doi:10.1360/TB-2020-1164 |
| [45] | Li Y X, Lei C Y, 1980. Fossil mammals from Lishui, Jiangsu. Vert PalAsiat, 18: 59-64 |
| [46] | Li Z, Dong W, 2007. Mammalian fauna from the Lingjing Paleolithic site in Xuchang, Henan Province. Acta Anthrop Sin, 26: 345-360 |
| [47] | Liu J L, Jin F Q, 1982. Several theoretical issues related to the Chaoxian fauna (2). J Huainan Min Coll, 1: 26-32 |
| [48] | Liu J Y, 1999. Mammal fossil remains of Late Pleistocene from Miaofengshan, Beijing, with a discussion on the dispersal and extinction of Crocuta C. ultima. Longgupo Prehist Cult, 1: 128-139 |
| [49] | Liu J Y, 2001. A review of Pachycrocuta sinensis and analysis of its relationship with the two extant genera Hyaena and Crocuta. In: Deng T, Wang Y eds. Proceedings of the Eighth Annual Meeting of the Chinese Society of Vertebrate Paleontology. Beijing: Ocean Press. 149-157 |
| [50] | Liu J Y, 2002. Section 4 Carnivora of Chapter 3 Mammals. In: Wu R K, Li X X, Wu X Z et al. eds. Homo erectus from Nanjing. Nanjing: Science and Technology Press, Jiangsu. 102-111 |
| [51] | Liu J Y in press. The Hyaenidae. In: Qiu Z X, Wang X M, Liu J Y eds. Creodonts and Carnivora - Creodonta and Carnivora: Feliformia. Palaeovertebrata Sinica. Vol. 7(1) |
| [52] | Liu J Y, Qiu Z X, 2009. Mammalian-Carnivora. In: Jin C Z, Liu J Y eds. Palaeolithic Site-The Renzidong Cave, Fanchang, Anhui Province. Beijing: Science Press. 220-282 |
| [53] | Liu J Y, Zheng L T, Xu Q Q et al., 2006. Study on carnivora fossil remains from the Jinpendong Cave, Wuhu, Anhui. Acta Anthrop Sin, 25: 227-241 |
| [54] | Liu J Y, Fang Y S, Zhang Z H, 2007. Carnivora. In: Nanjing Museum, Jiangsu Institute of Archaeology eds. The Early Pleistocene Mammalian Fauna at Tuozi Cave, Nanjing, China. Beijing: Science Press. 25-68 |
| [55] | Liu J Y, Zhao L X, Chen J et al., 2011. The age and environment study of Gigantopithecus fauna of Pa’eryan, Bijie, Guizhou, based on the carnivore fossils. Quat Sci, 31(4): 654-666 |
| [56] | Liu J Y, Wagner J, Chen P F et al., 2015. Mass mortality of a large population of the spotted hyenas (Crocuta ultima) at the Lingxiandong Cave, Qinhuangdao, Hebei Province: a hyena communal den with its palaeoecological and taphonomical interpretation. Quat Sci, 35(3): 607-621 |
| [57] | Liu J Y, Liu J Y, Zhang H W et al., 2021. The giant short-faced hyena Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae) from Northeast Asia: a reinterpretation of subspecies differentiation and intercontinental dispersal. Quat Int, 577: 29-51 |
| [58] | Liu J Y, Zhang Y Q, Chi Z Q et al., 2022. A Late Pliocene Hipparion houfenense fauna from Yegou, Nihewan Basin and its biostratigraphic significance. Vert PalAsiat, 60: 278-323 |
| [59] | Liu W H, Dong W, Zhang L M et al., 2017. New material of Early Pleistocene Sus (Artiodactyla, Mammalia) from Yangshuizhan in Nihewan Basin, North China. Quat Int, 434A: 32-47 |
| [60] | Ma A C, Tang H L, 1992. On discovery and significance of a Holocene Ailuropoda-Stegodon fauna from Jinhua, Zhejiang. Vert PalAsiat, 30: 295-312 |
| [61] | Mills G, 1982. Hyaena brunnea. Mamm Spec, 194: 1-5 |
| [62] | Mills G, 1990. Kalahari Hyaenas-Comparative Behavioral Ecology of Two Species. London: Unwin Hyman. 1-304 |
| [63] | Morales J, Senut B, Pickford M, 2011. Crocuta dietrichi from Meob, Namibia: implications for the age of the Tsondab Sandstone in the coastal part of the Namib Desert. Estud Geol, 67(2): 207-215 |
| [64] | Owen R, 1870. On fossil remains of mammals found in China. Quart J Geol Soc London, 26: 417-434 |
| [65] | Palmqvist P, Arribas A, 2001. Taphonomic decoding of the paleobiological information locked in a lower Pleistocene assemblage of large mammals. Paleobiology, 27: 512-530 |
| [66] | Palmqvist P, Martínez-Navarro B, Pérez-Claros J A et al., 2011. The giant hyena Pachycrocuta brevirostris: modelling the bone-cracking behavior of an extinct carnivore. Quat Int, 243: 61-79 |
| [67] | Pei J X, 1989. Thermolluminecence dating of the Peking Man Site and other caves. In: Wu R K, Wu X Z, Zhang S S eds. Early Humankind in China. Beijing: Science Press. 256-260 |
| [68] | Pei W C, 1934. On the carnivora from Locality 1 of Choukoutien. Palaeont Sin, Ser C, 8: 1-217 |
| [69] | Pei W C, 1939. Note on a collection of mammal fossils from Tanyang in Kiangsu Province. Bull Geol Soc China, 19(4): 379-392 |
| [70] | Pei W C, 1940. The Upper Cave fauna of Choukoutien. Palaeont Sin, New Ser C, 10: 1-84 |
| [71] | Pei W C, 1987. Carnivoira, Proboscidae and Rodentia from Liucheng Gigantopithecus cave and other caves in Guangxi. Mem Inst Vert Paleont Paleoanthrop Acad Sin, 18: 1-119 |
| [72] | Pei W C, Qiu Z L, 1957. On a collection of mammalian fossils from Liuhsia, Hongchow, Chekiang, China. Vert PalAsiat, 1: 42-46 |
| [73] | Pérez-Claros J A, 2024. Unravelling the origin of the brown hyena (Parahyaena brunnea) and its evolutionary and paleoecological implications for the Pachycrocuta. Palaeont Electron: 27.1.a18 |
| [74] | Petter G, Howell F C, 1989. Une nouvelle espèce du genre Crocuta Kaup (Mammalia: Carnivora: Hyaenidae) dans la faune pliocéne de Laetoli (Tanzanie): Crocuta dietrichi nov. sp.; origine du genre. C R Acad Sci, Paris, 308: 1031-1038 |
| [75] | Pilgrim G, 1931. Catalogue of the Pontian Carnivora of Europe in the Department of Geology. London: British Museum (Natural History). 1-174 |
| [76] | Qi G Q, 1989. Quaternary mammalian faunas and environment of fossil humans in Nouth China. In: Wu R K, Wu X Z, Zhang S S eds. Early Humankind in China. Beijing: Science Press. 277-337 |
| [77] | Qiao Y S, Guo Z T, Hao Q Z et al., 2002. Magnetostratigraphy and paleoclimatic significance of an eollian sequence from the Xuancheng area, Anhui Province. J Geom Echanics, 8(4): 369-375 |
| [78] | Qiu Z X, 1987. Die Hyaeniden aus dem Ruscinium und Villafranchium Chinas. Münchn Geowiss Abh A, 9: 1-108 |
| [79] | Qiu Z X, Deng T, Wang B Y, 2004. Early Pleistocene mammalian fauna from Longdan, Dongxiang, Gansu, Chian. Palaeont Sin, New Ser C, 27: 1-198 |
| [80] | Qiu Z X, Tedford R H in press. The Hyaenidae. In: Tedford R H, Qiu Z X, Flynn L J eds. Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals. Heidelberg: Springer |
| [81] | Rieger I, 1981. Hyaena hyaena. Mamm Spec, 50: 1-5 |
| [82] | Shen G J, Jin L H, 1992. U-series dating of speleothem samples from Guanyindong Cave, Qianxi county, Guizhou province. Acta Anthrop Sin, 11: 93-100 |
| [83] | Shen G J, Gao X, Gao B et al., 2009. Age of Zhoukoudian Homo erectus determined with 26Al/10Be burial dating. Nature, 458: 198-200 |
| [84] | Sheng G L, Soubrier J, Liu J Y et al., 2014. Pleistocene Chinese cave hyenas and the recent Eurasian history of the spotted hyena, Crocuta crocuta. Mole Ecol, 23(3): 522-533 |
| [85] | Simpson G, 1941. Large Pleistocene felines of North America. Am Mus Novit, 1136: 1-127 |
| [86] | Suraprasit K, Jaeger J J, Chaimanee Y et al., 2015. A complete skull of Crocuta crocuta ultima indicates a late Middle Pleistocene age for the Khok Sung (northeastern Thailand) vertebrate fauna. Quat Int, 374: 34-45 |
| [87] | Tomida Y, Jin C Z, 2009. Two new species of Pliopentalagus (Leporidae, Lagomorpha) from the Pliocene of Anhui Province, China, with a revision of Pl. huainanensis. Vert PalAsiat, 47: 53-71 |
| [88] | Tong H W, Wu X J, Dong Z et al., 2018. Preliminary report on the mammalian fossils from the ancient human site of Hualong Cave in Dongzhi, Anhui. Acta Anthrop Sin, 37: 284-305 |
| [89] | Tseng Z J, Chang C H, 2007. A study of new material of Crocuta crocuta ultima (Carnivora: Hyaenidae) from the Quaternary of Taiwan. Coll Res, 20: 9-19 |
| [90] | Tseng Z J, Jin C Z, Liu J Y et al., 2008. Fossil Hyaenidae (Mammalia: Carnivora) from Huainan, Anhui Province, China. Vert PalAsiat, 46: 133-146 |
| [91] | Turner A, 1984. The interpretation of variation in fossil specimens of spotted hyaena (Crocuta crocuta Erxleben, 1777) from Sterkfontein Valley sites (Mammalia: Carnivora). Ann Trans muse, 33(27): 399-418 |
| [92] | Turner A, 1990. The evolution of the guild of larger terrestrial carnivores during the Plio-Pleistocene in Africa. Geobios, 23: 349-368 |
| [93] | Turner A, 2001. Remains of Pachycrocuta brevirostris (Mammalia, Hyaenidae) from the lower Pleistocene site of Untermassfeld near Meiningen. In: Kahlke H D, Kahlke R D eds. Das Pleistozän von Untermassfeld in Thuringen. Bonn: Rudolf Harbelt Verlag. 673-690 |
| [94] | Turner A, Antón M, 1996. The giant hyena, Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae). Geobios, 29: 455-468 |
| [95] | Turner A, Antón M, Werdelin L, 2008. Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe. Geobios, 41: 677-687 |
| [96] | Valkenburgh B V, Ruff C B, 1987. Canine strength and killing behavior in larger carnivores. J Zool Lond, 212: 379-397 |
| [97] | Viret J, 1954. Le loess à bancs durcis de Saint-Vallier (Drôme), et sa faune de mammifères villafranchiens. Nouv Arch Mus Hist Nat, 4: 1-200 |
| [98] | Vislobokova I A, Agadjanian A K, 2015. New data on large mammals of the Pleistocene Trlica fauna, Montenegro, the central Balkans. Paleontol J, 49: 651-667 |
| [99] | Wang H, Li Z Y, Tong H W et al., 2022. Hominin paleoenvironment in East Asia: the Middle Paleolithic Xuchang-Lingjing (China) mammalian evidence. Quat Int, 633: 118-133 |
| [100] | Wang X M, Qiu Z X, in press. The introduction of Carnivora. In: Qiu Z X, Wang X M, Liu J Y et al. eds. Creodonts and Carnivora - Creodonta and Carnivora: Feliformia. Palaeovertebrata Sinica. Vol. 7(1) |
| [101] | Werdelin L, 1989. Constraint and adaptation in the bone-cracking canid Osteoborus (Mammalia: Canidae). Paleobiology, 15: 387-401 |
| [102] | Werdelin L, Lewis M E, 2012. The taxonomic identity of the type specimen of Crocuta sivalensis (Falconer, 1867). J Vert Paleont, 32(6): 1453-1456 |
| [103] | Werdelin L, Solounias N, 1991. The Hyaenidae: taxonomy, systematics and evolution. Fossils Strata, 30: 1-104 |
| [104] | Werdelin L, Solounias N, 1996. The evolutionary history of hyenas in Europe and western Asia during the Miocene. In: Bernor R L, Fahlbusch V, Rietschel S eds. Later Neogene European Biotic Evolution and Stratigraphic Correlation. New York: Columbia University Press. 290-306 |
| [105] | Westbury M V, Hartmann S, Barlow A et al., 2020. Hyena paleogenomes reveal a complex evolutionary history of cross-continental gene flow between spotted and cave hyena. Sci Adv, 6: eaay0456 |
| [106] | Xu C H, Zhang Y Y, Chen D D et al., 1984. Human occipital bone and mammalian fossils from Chaoxian, Anhui. Acta Anthrop Sin, 3: 202-209 |
| [107] | Xu Q Q, Zhang H, Xu H K, 1997. Mammalian fossils of Baiyundong Cave, Xuzhou and Loc.1 at Tangshan, Nanjing. Vert PalAsiat, 35: 217-223 |
| [108] | Ye Z Y, Yan D F, 1975. A new locality of Quaternary mammal fossils in Tongshan, Anhui Province. Vert PalAsiat, 13: 194-195 |
| [109] | You Y Z, Cai B Q, 1996. Stratigraphic division, fossil mammals and environment of the Pleistocene in Fujian Province. Acta Anthrop Sin, 15: 335-346 |
| [110] | You Y Z, Xu S B, 1996. Quaternary stratigraphy and mammal fossils of Yihe and Shuhe river drainages. Vert PalAsiat, 34: 322-331 |
| [111] | Zdansky O, 1927. Weitere Bemerkungen liber fossile Carnivoren aus China. Palaeont Sin, Ser C, 4(4): 1-28 |
| [112] | Zhang Z Q, 2001. Fossil mammals of Early Pleistocene from Ningyang, Shandong Proivince. Vert PalAsiat, 39: 139-150 |
| [113] | Zheng L T, 2004. “The Huaihe Transitional Zone” and Quaternary mammals in Anhui. Acta Anthrop Sin, 23(suppl): 315-324 |
| [114] | Zou S L, Chen X, Zhang B et al., 2016. Preliminary report on the Late Pleistocene mammalian fauna from Shangli County, Pingxiang, Jiangxi Province. Acta Anthrop Sin, 35: 109-120 |
| [1] | XU Xing-Dong, SHI Qin-Qin. First discovery of the spiral-horned antelope Antilospira (Bovidae, Artiodactyla) from the Linxia Basin, Gansu, China . Vertebrata Palasiatica, 2026, 64(1): 47-58. |
| [2] | WANG Shi-Qi, MA Jiao, FU Jiao, BAI Wei-Peng. A new cranium of Metacervocerus longdanensis (Cervidae, Mammalia) and the genus Metacervocerus in China . Vertebrata Palasiatica, 2026, 64(1): 59-73. |
| [3] | CHANG Mei-Jing, SHI Qin-Qin, NI Xi-Jun, LI Qiang. New material of Eospalax simplicidens (Spalacidae, Rodentia) from Jianyucha, Zichang, northern Shaanxi . Vertebrata Palasiatica, 2026, 64(1): 26-46. |
| [4] | JIANGZUO Qi-Gao, LI Hong, YAMAGUCHI Nobuyuki, Joan MADURELL-MALAPEIRA, ZHANG Jian-Sheng, MA Hui-Min, GUO Ding-Ge, LI Shi-Jie, FU Jiao, ZHANG Xiao-Xiao, LI Chun-Xiao, XIE Kun, TONG Hao-Wen, LIU Jin-Yi, WANG Shi-Qi, DENG Tao. First discovery of Panthera spelaea cranium from Salawusu, northern China . Vertebrata Palasiatica, 2025, 63(4): 323-334. |
| [5] | Rana Mehroz FAZAL, HE Zhan-Wu, DU Bao-Pu, CHANG Mei-Jing, SHI Jing-Song, NI Xi-Jun, LI Qiang, ZHAO Ling-Xia. Discovery of Pleistocene fruit bat Rousettus and its coexisting non-volant micromammalian fossils from the Guilin Basin, South China . Vertebrata Palasiatica, 2025, 63(3): 173-188. |
| [6] | TONG Hao-Wen, CHEN Xi, ZHANG Bei, SUN Ji-Jia. New fossil materials of Sus lydekkeri from the Early Pleistocene Shanshenmiaozui site in Nihewan Basin of North China . Vertebrata Palasiatica, 2025, 63(3): 210-241. |
| [7] | DONG Wei, LIU Wen-Hui, BAI Wei-Peng, LIU Si-Zhao, WANG Yuan, LIU Jin-Yuan, JIN Chang-Zhu. A skull of Early Pleistocene Paracamelus gigas (Artiodactyla, Mammalia) from Luotuo Hill in Dalian, Northeast China . Vertebrata Palasiatica, 2024, 62(1): 47-68. |
| [8] | DONG Wei, BAI Wei-Peng, LIU Wen-Hui, ZHANG Li-Min. The first description of Equidae (Perissodactyla, Mammalia) from Xinyaozi Ravine in Shanxi, North China . Vertebrata Palasiatica, 2023, 61(3): 212-244. |
| [9] | TONG Hao-Wen, ZHANG Bei, CHEN Xi, WANG Xiao-Min. New fossils of small and medium-sized bovids from the Early Site of Shanshenmiaozui in Nihewan Basin, North China . Vertebrata Palasiatica, 2022, 60(2): 134-168. |
| [10] | DONG Wei, BAI Wei-Peng, ZHANG Li-Min. The first description of Rhinocerotidae (Perissodactyla, Mammalia) from Xinyaozi Ravine in Shanxi, North China . Vertebrata Palasiatica, 2021, 59(4): 273-294. |
| [11] | SHEN Wei, Thomas A. STIDHAM, LI Zhi-Heng. Reexamination of the oldest pigeon (Aves: Columbidae) from Asia: Columba congi from the Early Pleistocene of Zhoukoudian, Beijing, China . Vertebrata Palasiatica, 2021, 59(3): 245-256. |
| [12] | SUN Ji-Jia, ZHANG Bei, CHEN Xi, DENG Li, WEN Jun, TONG Hao-Wen. New fossils of Late Pleistocene Sus scrofa from Yangjiawan Cave 2, Jiangxi, China . Vertebrata Palasiatica, 2021, 59(1): 64-80. |
| [13] | DONG Wei, BAI Wei-Peng, PAN Yue, LIU Wen-Hui. New material of Cervidae (Artiodactyla, Mammalia) from Xinyaozi Ravine in Shanxi, North China . Vertebrata Palasiatica, 2020, 58(3): 221-248. |
| [14] | DONG Wei, LIU Wen-Hui, BAI Wei-Peng. Cladistic approach on chronological relationship of the Pleistocene mammalian faunas from China . Vertebrata Palasiatica, 2020, 58(1): 67-81. |
| [15] | ZHENG Shao-Hua, ZHANG Ying-Qi, CUI Ning. Five new species of Arvicolinae and Myospalacinae from the Late Pliocene-Early Pleistocene of Nihewan Basin . Vertebrata Palasiatica, 2019, 57(4): 308-324. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||