New fossils of small and medium-sized bovids from the Early Site of Shanshenmiaozui in Nihewan Basin, North China
TONG Hao-Wen, ZHANG Bei, CHEN Xi, WANG Xiao-Min
Vertebrata Palasiatica
2022, 60 ( 2):
134-168.
DOI: 10.19615/j.cnki.2096-9899.220413
Shanshenmiaozui site in Nihewan Basin in North China is a recently discovered Early Pleistocene site which yields rich and diverse mammalian fossils. In the fauna, the small and medium-sized bovid fossils are well represented and can be referred to the following taxa: Spirocerus wongi, Gazella sinensis, Ovis shantungensis and Megalovis piveteaui respectively, among which G. sinensis is the dominate species. S. wongi and G. sinensis are mainly represented by horn-cores and partial skull bones as well as mandibles; in addition, metacarpal and/or metatarsal bones were also recognized for all of the four species. The horn-cores are easy to be identified to the species level, while the dentitions and the postcranial bones underwent a series of examinations and comparisons before getting properly determined and referred to the most approximate taxa. Among the postcranial bones, the metapodials, especially to the metacarpal bones special attentions were paid, which are crucial not only for taxonomic identification, but also for phylogenetic and paleoecological reconstructions; the previously misidentified metapodial specimens in Nihewan fauna were reconsidered in this paper. In the SSMZ fauna, the bovid guild is dominated by Gazella and Bison, which indicates steppe was the most important biome in Nihewan Basin during Early Pleistocene.
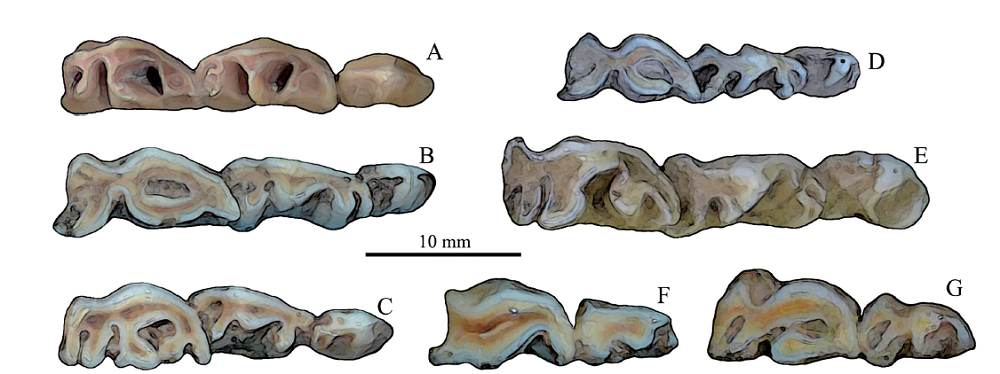
Fig. 8
Comparison of premolar series among some Quaternary gazelle species of China
A. Gazella sinensis, left p2-4, IVPP V 28681.2, SSMZ; B. Procapra gutturosa, left p2-4, NWIPB 0006065, extant; C. P. przewalskii, right p2-4 (horizontally flipped), NWIPB 0001172, extant;D. P. picticaudata, left p2-4, NWIPB 0001179, extant; E. Gazella subgutturosa, left p2-4, IVPP-c-05, extant;F. Saiga tatarica, right p3-4 (horizontally flipped), NWIPB S-80503, extant;G. Pantholops hodgsonii, left p3-4, NWIPB 77001, extant. All are in occlusal views
Extracts from the Article
Comparisons and discussions Premolars are crucial for taxonomic identification for the gazelles, especially the tooth structures of the lower p4 which is usually regarded as the most practical tooth in taxonomic distinctions (Boule et al., 1928; Teilhard de Chardin and Piveteau, 1930; Gentry, 1966; Janis and Lister, 1985; Chen, 1997). Among all the Gazella species, including both extant and fossil forms, the lower p4 usually has open lingual valleys (Teilhard de Chardin and Piveteau, 1930; Teilhard de Chardin and Trassaert, 1938; Zhang and Yang, 2016; Li et al., 2018) (Fig. 7M, 8E), except G. sinensis (Fig. 7C4, D3, F-J; 8A) and G. yushensis Chen, 1997; whereas the Capra species usually has the anterior valley closed in p4, i.e. paraconid and metaconid joined to form a continuous anterolingual wall (Fig. 7O), and the posterior valley sometimes is also closed. Among all the Gazella species, G. sinensis has the most complicated lower p3, some of which have the anterior valleys partially closed lower in crown, and all the cuspids and stylids are well developed, especially the parastylid is very robust, and the metaconid is the most developed relative to other compared Asian gazelles’ (Fig. 8A); but most p3s have their lingual valleys open. Because of the preorbital fossa, the fossils represented by G. sinensis were included into the genus Gazella; on the other hand, its lower p4 has closed anterior valley, which is a crucial character of Procapra; therefore, the species G. sinensis was considered as the direct ancestor of Procapra (Sokolov and Lushchekina, 1997).
In addition to lower p4, there exist other crucial premolar characters to distinguish the gazelle taxa in China, e.g. Saiga tatarica usually lacks and Pantholops hodgsonii exclusively lacks p2 in their dentitions (Fig. 8F-G).
# Calculated by the first author of this paper; mean values in parentheses. ... The history of the origin, evolution and dispersal of the Late Pleistocene Mammuthus-Coelodonta faunal complex in Eurasia (large mammals) 1 1999 ... Spirocerus was established by Boule and Teilhard de Chardin in 1928, and included the specimens from Sjara-osso-gol (=Salawusu) of Nei Mongol (Inner Mongolia) to it. Later on, the similar fossils from Nihewan (Teilhard de Chardin and Piveteau, 1930) and Yushe (Teilhard de Chardin and Trassaert, 1938) also were referred to this genus. Spirocerus was originally included into the subfamily Tragelaphinae by Teilhard de Chardin and Trassaert (1938), later on, it was referred to the subfamily Caprine (Sokolov, 1953); while currently it is usually placed in the subfamily Antilopinae (McKenna and Bell, 1997; http://taxonomicon.taxonomy.nl/TaxonTree.aspx?src=0&id=68446). More recent assignment is to Bovinae (Bai et al., 2019). Up to now, the classifications of Spirocerus at the species level are mainly based on horn-core characters, which caused quite a lot of confusions. Kahlke (1999) proposed that the most primitive Spirocerus with only anterior carena should be referred to S. wongi. Concerning the bicarinate forms with longer and slender horn-cores, the stratigraphically older ones can be referred to S. kiakhtensis peii, while the younger materials with correspondingly shorter and more massive horn-cores to S. kiakhtensis kiakhtensis. Moreover, all of the species of Spirocerus have heteronymous spiraled horns. ... Revision of some Late Miocene spiral horned antelopes (Bovidae, Mammalia) 1 2004 ... Comparison and discussion The degree and direction of horn-core torsion, the type of spiraling and the development and position of the keels are the crucial characters to classify the fossil spiral-horned antelopes (Kostopoulos, 2004). In the early publications, the torsion direction (clockwise or anticlockwise) always means for the right horn (Pilgrim, 1939). Moreover, the recent publications prefer to use “homonymous” and “heteronymous”, if the left horn twists clockwise, the horns are termed homonymous, or else it’s heteronymous. ... Revision of phylogenetic relationships in the Antilopinae Subfamily on the basis of the mitochondrial rRNA and β-Spectrin nuclear gene sequences 1 2003 ... The suprageneric classification of Bovidae is after Pilgrim (1939), Sokolov (1953), Gentry (1992, 2010), McKenna and Bell (1997), Vrba and Schaller (2000), Grubb (2001), Matthee and Davis (2001), Kuznetsova and Kholodova (2003), Hassanin et al. (2012); the taxonomy at generic level is after Groves (2014) and Castelló (2016); the classification of the local Antilopinae and Caprinae is after Jiang (2004). ... “Gazella” (Mammalia: Bovidae) from the Late Miocene Qingyang area, Gansu, China 1 2018 ... Comparisons and discussions Premolars are crucial for taxonomic identification for the gazelles, especially the tooth structures of the lower p4 which is usually regarded as the most practical tooth in taxonomic distinctions (Boule et al., 1928; Teilhard de Chardin and Piveteau, 1930; Gentry, 1966; Janis and Lister, 1985; Chen, 1997). Among all the Gazella species, including both extant and fossil forms, the lower p4 usually has open lingual valleys (Teilhard de Chardin and Piveteau, 1930; Teilhard de Chardin and Trassaert, 1938; Zhang and Yang, 2016; Li et al., 2018) (Fig. 7M, 8E), except G. sinensis (Fig. 7C4, D3, F-J; 8A) and G. yushensis Chen, 1997; whereas the Capra species usually has the anterior valley closed in p4, i.e. paraconid and metaconid joined to form a continuous anterolingual wall (Fig. 7O), and the posterior valley sometimes is also closed. Among all the Gazella species, G. sinensis has the most complicated lower p3, some of which have the anterior valleys partially closed lower in crown, and all the cuspids and stylids are well developed, especially the parastylid is very robust, and the metaconid is the most developed relative to other compared Asian gazelles’ (Fig. 8A); but most p3s have their lingual valleys open. Because of the preorbital fossa, the fossils represented by G. sinensis were included into the genus Gazella; on the other hand, its lower p4 has closed anterior valley, which is a crucial character of Procapra; therefore, the species G. sinensis was considered as the direct ancestor of Procapra (Sokolov and Lushchekina, 1997). ... The Pliocene lacustrine series in central Shansi 1 1935 ... In China, Nihewan Basin and Yushe Basin are the most important fossiliferous spots for Late Cenozoic bovids, but the latter’s bovid fauna is much more diversified (Teilhard de Chardin and Trassaert, 1938) than the former’s. In Yushe Basin of Shanxi Province, the late Cenozoic deposits were subdivided into three biostratigraphical zones from bottom to top: Zone I, Zone II and Zone III (Licent and Trassaert, 1935; Teilhard de Chardin and Trassaert, 1938; Tedford et al., 1991); the Zone III shares almost all of its faunal members with the Nihewan fauna. In regard to the bovid taxa, only the advanced and relict ones of each group of Yushe Basin also appear in the Nihewan Basin; the relict elements include Gazella, Antilospira and Megalovis; the newly appeared taxa include Spirocerus, Ovis and Bison. The recently discovered fauna in Jinyuan Cave in Dalian of Liaoning Province bears a very similar bovid assemblage (Jin et al., 2021) with the Nihewan fauna. ... The morphology of the lower fourth premolar as a taxonomic character in the Ruminantia (Mammalia; Artiodactyla), and the systematic position of Triceromeryx 1 1985 ... Lower teeth: The lower premolars are less hypsodont than those of the caprinines, but complicatedly constructed, except p2 which is simply constructed and prominently smaller than the other two (Table 6). Lower p3 and p4 has the similar structure, both of them are elongated but p3 has the lingual wall open both mesially and distally (Fig. 7C4, D1, G-H), sometimes with the anterior valley partially closed (Figs. 7D1; 8A); on the other hand, the p4 only has the posterior valley open, like a narrow groove (Figs. 7C4, D3, F-J; 8A) and coincides with the type 5 of p4 patterns proposed by Janis and Lister (1985). Each premolar has two roots. Lower molars are elongate and with developed goat folds, but lack basal pillars; m1 and m2 are very similar in size and shape; m3 has three lobes, but the third lobe has very shallow infundibulum. Molars are quite hypsodont. The m1 and m2 with two roots, but m3 has three roots. ... Magnetostratigraphic dating of the Xiashagou Fauna and implication for sequencing the mammalian faunas in the Nihewan Basin, North China 2 2012 ... The SSMZ site lies at the neighboring hill of Xiaochangliang, a well-known Paleolithic Site in Nihewan Basin. Based on the stratigraphic correlation in the field, the fossil-bearing sand-silt bed at SSMZ site is a little higher than the cultural layer at XCL site (Tong et al., 2011; Liu et al., 2012), whose paleomagnetic age is about 1.36 Ma BP (Zhu et al., 2001). On the other hand, the SSMZ fauna is very similar with the classical Nihewan fauna in faunal composition, which means the SSMZ fauna should have an approximate age as the latter based on faunal correlation (Tong et al., 2021), whose recent dating result is 2.2-1.7 Ma (Liu et al., 2012). ...
Other Images/Table from this Article
-
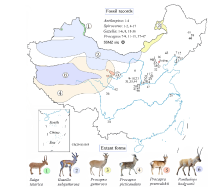 Fig. 1
Location of Shanshenmiaozui (SSMZ) site, with distributional map of living antelopes and related fossil forms of the Pleistocene Epoch in China Data for the extant taxa are from Jiang (2004), Smith and Xie (2008). Fossil localities: 1. Xiashagou of Nihewan; 2. Yushe; 3. Dongdoubi of Yuxian; 4. Jinyuan Cave; 5. Zhoukoudian Loc.1;6. Xujiayao; 7. Mashandong; 8. Salawusu; 9. Xifeng of Qingyang; 10. Loufangzi; 11. Jinniushan;12. Miaohoushan; 13. Haimao of Dalian; 14. Gulongshan; 15. Xiaogushan; 16. Chicheng; 17. Dingcun;18. Xihoudu; 19. Tianshuigou of Dali; 20. Wenxi; 21. Pinglu; 22. Danangou of Yuxian; 23. Bajiazui;24. Gonghe; 25. Yangguo; 26. Tuozidong; 27. Tunliu; 28. Banpo; 29. Linyi; 30. Yuanmou; 31. Heshui;32. Xingtai; 33. Xiaochangcun; 34. Wuwang of Linyi; 35. Gengjiagou; 36. Luhuo; 37. Zhoukoudian Loc.15;38. Upper Cave; 39. Guxiangtun; 40. Zhoujiayoufang; 41. Dali Man site; 42. Aba; 43. Chifeng;44. Rouyuan; 45. Yanjiagang; 46. Yuhongcun of Dali; 47. Lingjing
Fig. 1
Location of Shanshenmiaozui (SSMZ) site, with distributional map of living antelopes and related fossil forms of the Pleistocene Epoch in China Data for the extant taxa are from Jiang (2004), Smith and Xie (2008). Fossil localities: 1. Xiashagou of Nihewan; 2. Yushe; 3. Dongdoubi of Yuxian; 4. Jinyuan Cave; 5. Zhoukoudian Loc.1;6. Xujiayao; 7. Mashandong; 8. Salawusu; 9. Xifeng of Qingyang; 10. Loufangzi; 11. Jinniushan;12. Miaohoushan; 13. Haimao of Dalian; 14. Gulongshan; 15. Xiaogushan; 16. Chicheng; 17. Dingcun;18. Xihoudu; 19. Tianshuigou of Dali; 20. Wenxi; 21. Pinglu; 22. Danangou of Yuxian; 23. Bajiazui;24. Gonghe; 25. Yangguo; 26. Tuozidong; 27. Tunliu; 28. Banpo; 29. Linyi; 30. Yuanmou; 31. Heshui;32. Xingtai; 33. Xiaochangcun; 34. Wuwang of Linyi; 35. Gengjiagou; 36. Luhuo; 37. Zhoukoudian Loc.15;38. Upper Cave; 39. Guxiangtun; 40. Zhoujiayoufang; 41. Dali Man site; 42. Aba; 43. Chifeng;44. Rouyuan; 45. Yanjiagang; 46. Yuhongcun of Dali; 47. Lingjing
-
 Table 1
Studied fossil specimens of small to medium-sized bovids newly unearthed from SSMZ
Table 1
Studied fossil specimens of small to medium-sized bovids newly unearthed from SSMZ
-
 Fig. 2
Partial skulls, jaw bones and teeth of Spirocerus wongi (A-D) and Ovis shantungensis (E) from SSMZ A-D. Spirocerus wongi: A. partial skull with horn-cores (IVPP V 28650), B1-B3. juvenile maxilla with DP2-M1 (V 28651), C1-C4. left mandible with dp2-m1 (V 28652.1), D1-D4. partial right mandible with p4-m3 (V 28653); E1-E2. Ovis shantungensis, maxilla with left DP4-M1 and right DP2-4 and M1 (V 28693) A. anterior view; B1, E1. palatal views; B2, C1, D1, E2. buccal views; C2, D2. lingual views;B3, C3-4, D3-4. occlusal views. The unmarked scale bars equal 20 mm
Fig. 2
Partial skulls, jaw bones and teeth of Spirocerus wongi (A-D) and Ovis shantungensis (E) from SSMZ A-D. Spirocerus wongi: A. partial skull with horn-cores (IVPP V 28650), B1-B3. juvenile maxilla with DP2-M1 (V 28651), C1-C4. left mandible with dp2-m1 (V 28652.1), D1-D4. partial right mandible with p4-m3 (V 28653); E1-E2. Ovis shantungensis, maxilla with left DP4-M1 and right DP2-4 and M1 (V 28693) A. anterior view; B1, E1. palatal views; B2, C1, D1, E2. buccal views; C2, D2. lingual views;B3, C3-4, D3-4. occlusal views. The unmarked scale bars equal 20 mm
-
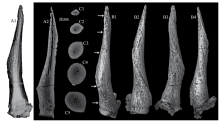 Fig. 3
CT scan images and 3-D reconstructions of the horn-core of Spirocerus wongi from SSMZ (IVPP V 28650) A1-A2. CT scan images showing the general canal system (A1) and a longitudinal slice (A2);C1-C5. CT scan slices showing the changes of cross sections and the canal system at different levels;B1-B4. CT image reconstruction of the right horn-core in anterior (B1), lateral (B2), posterior (B3) and medial (B4) views
Fig. 3
CT scan images and 3-D reconstructions of the horn-core of Spirocerus wongi from SSMZ (IVPP V 28650) A1-A2. CT scan images showing the general canal system (A1) and a longitudinal slice (A2);C1-C5. CT scan slices showing the changes of cross sections and the canal system at different levels;B1-B4. CT image reconstruction of the right horn-core in anterior (B1), lateral (B2), posterior (B3) and medial (B4) views
-
Table 2
Measurements of the horn-cores of Spirocerus wongi, compared with related species (mm)
-
Table 3
Measurements of the teeth of Spirocerus wongi, compared with related species (mm)
-
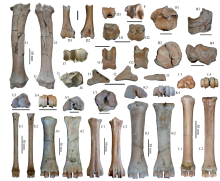 Fig. 4
Postcranial bones of small and medium-sized bovids from SSMZ A-C. Spirocerus wongi: A1-A3. left Mc III+IV (IVPP V 28655),B1-B4. right naviculo-cuboid+lat-mid+medial cuneiforms (V 28656.2-3),C1-C3. right Mt III+IV (V 28656.4); D-I. Gazella sinensis: D1-D3. partial left humerus (V 28687),E1-E4. left Mc III+IV (V 28688), F. distal epiphysis of left tibia (V 28691),G1-G2. left astragalus (V 28692), H1-H2. left naviculo-cuboid (V 28690), I1-I3. 3rd phalanx (V 28689);J-K. Megalovis piveteaui: J1-J4. left radius (V 28654), K1-K4. right Mc III+IV (V 28695);L1-L4. Ovis shantungensis: left Mc III+IV (V 28694). A1, C1, D1, E1, G1, J1, K1, L1. anterior views;A2, B4, C2, D2, E2, G2, J2, K2, L2. posterior views; A3, B1, C3, E3, H1, J3, K3, L3. proximal views;B2, D3, E4, F, H2, J4, K4, L4. distal views; B3. medial view; I1. lateral view; I2. interdigital view;I3. volar view. The arrows indicate the lateral projection of the radius. The unmarked scale bars equal 20 mm
Fig. 4
Postcranial bones of small and medium-sized bovids from SSMZ A-C. Spirocerus wongi: A1-A3. left Mc III+IV (IVPP V 28655),B1-B4. right naviculo-cuboid+lat-mid+medial cuneiforms (V 28656.2-3),C1-C3. right Mt III+IV (V 28656.4); D-I. Gazella sinensis: D1-D3. partial left humerus (V 28687),E1-E4. left Mc III+IV (V 28688), F. distal epiphysis of left tibia (V 28691),G1-G2. left astragalus (V 28692), H1-H2. left naviculo-cuboid (V 28690), I1-I3. 3rd phalanx (V 28689);J-K. Megalovis piveteaui: J1-J4. left radius (V 28654), K1-K4. right Mc III+IV (V 28695);L1-L4. Ovis shantungensis: left Mc III+IV (V 28694). A1, C1, D1, E1, G1, J1, K1, L1. anterior views;A2, B4, C2, D2, E2, G2, J2, K2, L2. posterior views; A3, B1, C3, E3, H1, J3, K3, L3. proximal views;B2, D3, E4, F, H2, J4, K4, L4. distal views; B3. medial view; I1. lateral view; I2. interdigital view;I3. volar view. The arrows indicate the lateral projection of the radius. The unmarked scale bars equal 20 mm
-
Table 4
Measurements of metacarpals of bovids from Nihewan Basin, compared with related taxa (mm)
-
 Fig. 5
Incomplete skulls and horn-cores of Gazella sinensis from SSMZ A. partial skull of a juvenile with horn-cores (IVPP V 28658); B1-B3. partial skull with horn-cores (V 28657); C1-C2. left and right horn-cores (V 28659.1, V 28659.2); D1-D2. partial skull with left horn-core (V 28667);E. partial skull with right horn-core (V 28661). A, B1, C1-2, D1. anterior views; B3. posterior view;B2. dorsal view; E. medial view; D2. lateral view. Scale bars equal 50 mm
Fig. 5
Incomplete skulls and horn-cores of Gazella sinensis from SSMZ A. partial skull of a juvenile with horn-cores (IVPP V 28658); B1-B3. partial skull with horn-cores (V 28657); C1-C2. left and right horn-cores (V 28659.1, V 28659.2); D1-D2. partial skull with left horn-core (V 28667);E. partial skull with right horn-core (V 28661). A, B1, C1-2, D1. anterior views; B3. posterior view;B2. dorsal view; E. medial view; D2. lateral view. Scale bars equal 50 mm
-
Table 5
Measurements of partial cranial bones and horn-cores of Gazella sinensis (mm)
-
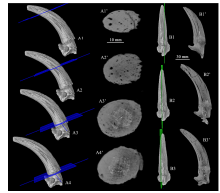 Fig. 6
CT scan images of the horn-core of Gazella sinensis (IVPP V 28661) from SSMZ A1-A4. CT image reconstruction showing positions of the cross CT scan slices (A1’-A4’);B1-B3. CT image reconstruction showing positions of the longitudinal CT scan slices (B1’-B3’)
Fig. 6
CT scan images of the horn-core of Gazella sinensis (IVPP V 28661) from SSMZ A1-A4. CT image reconstruction showing positions of the cross CT scan slices (A1’-A4’);B1-B3. CT image reconstruction showing positions of the longitudinal CT scan slices (B1’-B3’)
-
 Fig. 7
Jaw bones and teeth of Gazella sinensis from SSMZ (A-J), compared with related taxa (K-P) A. maxilla with P3-M1 (IVPP V 28673); B. maxilla with M1-3 (V 28674);C1-C4. left mandible with p2-m3 (V 28680); D1-D3. left mandible with p2-m3 (V 28681.2);E1-E2. left mandible with dp2-4 and m1-2 (V 28685.1), E3. detail of dp2-4; F. right p4-m3 (V 28677);G. left p3-m3 (V 28682); H. right p2-m3 (V 28675); I. left p4-m3 (V 28686); J. left p4-m3 (V 28678);K. Procapra przewalskii, right p2-m3 (horizontally flipped) (NWIPB-0001172♀);L. Procapra gutturosa, left p2-m3 (NWIPB 620032); M. Gazella subgutturosa, left p2-m3 (NWIPB 609001);N. Pseudois nahaur, left p2-m3 (NWIPB-KX1); O. Capra ibex, left p2-m3 (IOZ-2);P. Ovis ammon, left p2-m3 (OV 1346-2). A, B, C1, C4, D1, E3, F-P. occlusal views;C2, D2, E1. buccal views; C3, D3, E2. lingual views. The arrows indicate the variations of p4 The unmarked scale bars equal 20 mm
Fig. 7
Jaw bones and teeth of Gazella sinensis from SSMZ (A-J), compared with related taxa (K-P) A. maxilla with P3-M1 (IVPP V 28673); B. maxilla with M1-3 (V 28674);C1-C4. left mandible with p2-m3 (V 28680); D1-D3. left mandible with p2-m3 (V 28681.2);E1-E2. left mandible with dp2-4 and m1-2 (V 28685.1), E3. detail of dp2-4; F. right p4-m3 (V 28677);G. left p3-m3 (V 28682); H. right p2-m3 (V 28675); I. left p4-m3 (V 28686); J. left p4-m3 (V 28678);K. Procapra przewalskii, right p2-m3 (horizontally flipped) (NWIPB-0001172♀);L. Procapra gutturosa, left p2-m3 (NWIPB 620032); M. Gazella subgutturosa, left p2-m3 (NWIPB 609001);N. Pseudois nahaur, left p2-m3 (NWIPB-KX1); O. Capra ibex, left p2-m3 (IOZ-2);P. Ovis ammon, left p2-m3 (OV 1346-2). A, B, C1, C4, D1, E3, F-P. occlusal views;C2, D2, E1. buccal views; C3, D3, E2. lingual views. The arrows indicate the variations of p4 The unmarked scale bars equal 20 mm
-
Table 6
Measurements of teeth of Gazella sinensis from SSMZ, compared with those from Xiashagou (mm)
-
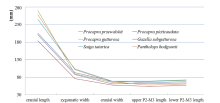 Fig. 9
Toothrow length and cranial size of extant gazelles in China Raw data was employed from Jiang, 2004
Fig. 9
Toothrow length and cranial size of extant gazelles in China Raw data was employed from Jiang, 2004
-
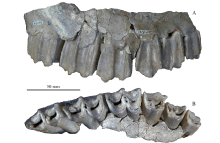 Fig. 10
Maxilla of Megalovis piveteaui (MNHN-NIH 150) from Xiashagou of Nihewan A. buccal view; B. occlusal view
Fig. 10
Maxilla of Megalovis piveteaui (MNHN-NIH 150) from Xiashagou of Nihewan A. buccal view; B. occlusal view
-
 Fig. 11
Length vs distal width of metacarpal bones of diverse bovids The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
Fig. 11
Length vs distal width of metacarpal bones of diverse bovids The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
|

{kind=link}