New fossils of small and medium-sized bovids from the Early Site of Shanshenmiaozui in Nihewan Basin, North China
TONG Hao-Wen, ZHANG Bei, CHEN Xi, WANG Xiao-Min
Vertebrata Palasiatica
2022, 60 ( 2):
134-168.
DOI: 10.19615/j.cnki.2096-9899.220413
Shanshenmiaozui site in Nihewan Basin in North China is a recently discovered Early Pleistocene site which yields rich and diverse mammalian fossils. In the fauna, the small and medium-sized bovid fossils are well represented and can be referred to the following taxa: Spirocerus wongi, Gazella sinensis, Ovis shantungensis and Megalovis piveteaui respectively, among which G. sinensis is the dominate species. S. wongi and G. sinensis are mainly represented by horn-cores and partial skull bones as well as mandibles; in addition, metacarpal and/or metatarsal bones were also recognized for all of the four species. The horn-cores are easy to be identified to the species level, while the dentitions and the postcranial bones underwent a series of examinations and comparisons before getting properly determined and referred to the most approximate taxa. Among the postcranial bones, the metapodials, especially to the metacarpal bones special attentions were paid, which are crucial not only for taxonomic identification, but also for phylogenetic and paleoecological reconstructions; the previously misidentified metapodial specimens in Nihewan fauna were reconsidered in this paper. In the SSMZ fauna, the bovid guild is dominated by Gazella and Bison, which indicates steppe was the most important biome in Nihewan Basin during Early Pleistocene.
| Mc III+IV | | Taxa | Sources | Length | Proximal width | Proximal APD | Distal width | Distal APD | Midshaft width | Midshaft APD | Index of stoutness (Distal width/total length) (%) | | Bison palaeosinensis | Teilhard and Piveteau (1930) | 231-248 | 72-74 | | 65 | | | | 28-29 | | Tong et al. (2018) | 226-233 | 60-67.8 | 35-38.6 | 59-63.5 | 34.2-34.7 | 36-39.5 | 26-27.6 | 26-28 | | Megalovis latifrons | Viret (1954) | 200 | 56.5 | | 60 | | | | 30 | | Megalovis latifrons | Radulesco and Samson (1962) | 192 | 56 | | 64 | | 35.7 | | 33.3 | | Megalovis piveteaui | This paper | 212 | 59 | 37 | 59 | 32 | 34 | 25 | 27.8 | | Teilhard and Piveteau (1930); Schaub (1937); Radulesco and Samson (1962) | 192-204 | 51-57 | | 54-61 | | | | 28-30# | | Megalovis wimani | Schaub (1937) | 185 | 52 | | 57 | | | | 30.8# | | Ovis shantungensis | This paper | 221 | 41 | 29 | 42 | 27 | 26 | 20 | 19 | | Teilhard and Piveteau (1930) | 231-233 | 42-44 | | 42-44.5 | | | | 18-19 | | Ovis ammon | Fedosenko and Blank (2005) | 218 | | | | | | | | | Ovis shangi | Teilhard and Young (1936) | 150 | 26 | | 28 | | 17 | | 18.7 | | Spirocerus wongi | This paper | 175 | 47 | 32 | 56 | 29 | 31 | 19 | 32 | | Spirocerus kiakhtensis | Sokolov (1959) | 170 | | | 60.9# | | 35.0# | 22.0# | 35.8 | | Gazella cf. G. lydekkeri | Zhang and Yang (2016) | 141 | | | | | | | | | Gazella sinensis | This paper | 180 | 22 | 17 | 23 | 16 | 12.5 | 11.3 | 12.8 | | Gazella subgutturosa | Demircioglu and Ince (2020) | 146.0(F) | 18.4(F) | | 18.0(F) | | 10.5(F) | 9.7(F) | | | | 150.7(M) | 19.6(M) | | 18.9(M) | | 11.4(M) | 9.9(M) | | | Procapra przewalskii | Boule et al. (1928) | 142 | | | 18-21 | | | | 12.7-14.8 | | Procapra gutturosa | Boule et al. (1928) | 180 | | | 23 | | | | 12.8 | | Procapra gutturosa | This paper (OV 1124) | 146 | 15.5 | 8.0 | 16.7 | 12.1 | 8.7 | 7.0 | 11.4 | | Saiga tatarica | Prat (1966b) | 153 | 25 | 18.5 | 25.5 | | 13.5 | | 17.7 | | Capra ibex | Prat (1966a) | 126-155 | 28.5-38.5 | 20.5-27 | 31.5-43 | 19-24 | 20-27.5 | | | | | (139.2) | (33) | (23.8) | (36.7) | (21.3) | (23.8) | | | | Praeovibos priscus | Kahlke (1964) | 193.7# | | | | | | | | | Ovibos moschatus sussenbornensis | Kahlke (1961) | 189.9# | 62.8# | 36.5# | 69.1# | | 44# | | 36.4 | | Mt III+IV | | Spirocerus wongi | This paper | 173.0 | 36.9 | 34.0 | 45.3 | 26.1 | 22.0 | 21.5 | 26.2 |
Table 4
Measurements of metacarpals of bovids from Nihewan Basin, compared with related taxa (mm)
Extracts from the Article
Mc III+IV: The Mc III+IV (Fig. 4A1-3) is a kind of short but robust one, with the distal end expanded. The cranial aspect is more rounded on the shaft while the caudal surface is more flat. At both the proximal and distal ends the medial side or Mc III looks prominently thicker than the lateral side. In proximal view, there are two large articular facets, the larger facet or magnum trapezoid facet is relatively wide and has a curved edge, which is articulated with the fused 2nd and 3rd carpals; the smaller facet or unciform facet is triangular and articulates with the 4th carpal; between the two facets there exists a groove or depression. The posterior medial tubercle is not developed. In anterior view, the vascular groove (or metacarpal gully) is nearly unobservable and the nutrient foramen is tiny. In posterior view, the shaft has a quite flat surface, except near the proximal end, there is a depression; a small nutrient foramen exists near the distal articular surface. The dimensions are shown in Table 4.
Mt III+IV (Fig. 4C1-3): In general view, the Mt III+IV has the very proximate length as the Mc III+IV, which is quite different from the common ungulates whose metatarsals are prominently longer than metacarpals belonging to the same individual; furthermore, the distal end expands more sharply relative to the shaft. In proximal view, the proximal surface has a polygon outline, and consists of four articular facets; the two anterior facets are large and kidney-shaped, but the two posterior facets are small and elongated. The mediolateral dimension of the proximal end is slightly larger than the anteroposterior dimension. The two large articular facets meet at the front middle part but diverge posteriorly; both of them have longer anteroposterior dimensions. The anteromedial facet articulates with the fused 2nd and 3rd tarsals; the anterolateral facet articulates with naviculo-cuboid. There are two smaller facets located near the posterior margin. The posterolateral facet has larger mediolateral dimension and articulates with naviculo-cuboid. The small roundish facet articulates with the 1st tarsal. The dimensions are shown in Table 4.
Mc III+IV: A complete left Mc III+IV (IVPP V 28688) (Fig. 4E1-4). In general, the bone is very slender; the cranial aspect is more rounded on the shaft and the caudal aspect is much flatter and slightly concave at the upper part; the distal epiphysis is not fused yet. In proximal view, there are two articular facets, the larger one or magnum trapezoid facet is prominently wider and deeper, which articulates with the fused 2nd and 3rd carpals; the smaller one or unciform facet is oval-like and is the articular surface for 4th carpal; between the two facets there exists a narrow ridge; no nutrient foramen can be observed at the proximal end (Fig. 4E3). In anterior view, a prominent tubercle (extensor carpi radialis insertion: Gentry, 1966: fig. 12) occurs at the cranial aspect of the proximal end; no vascular groove (or metacarpal gully) can be observed, while the nutrient foramen is clear but small and located near the epiphyseal suture; the widest portion lies at the epiphyseal suture rather than at the trochlea (Fig. 4E1). In posterior view, no prominent groove can be observed except the upper part; the articular facet for the vestigial 5th metacarpal is absent. A prominent nutrient foramen exists near the epiphyseal suture and a tiny nutrient foramen occurs at the mid-shaft (Fig. 4E2). The dimensions are shown in Table 4.
The metacarpal bone is prominently larger than such allied taxa, as Gazella subgutturosa and Procapra przewalskii, but with the same size as that of P. gutturosa (Table 4); in morphology, it has no peculiarity. The metapodial bone does have distinctive characters relative to those of the caprines in their extremely slenderness. Meanwhile, the BT (maximum breadth of trochlea) of the metacarpal of SSMZ specimen is 20.9 mm, which stands between those of the Gazella (15.1-20.1) and Procapra (21.5-21.6) (Wang et al., 2020).
Metacarpal: A right anterior canon bone (Mc III+IV) (IVPP V 28695) (Fig. 4K1-4). In general, both the proximal and distal ends expand to some extent, and the metacarpal bone is moderately stout. At both proximal and anterior aspects, the medial side or Mc III looks noticeably thicker than the lateral side. The two proximal articular facets are quite flat, and a narrow groove occurs between the facets. In anterior view, the shaft is roundish, and the vascular groove (or metacarpal gully) is very narrow at the upper par but widened around the nutrient foramen. In posterior view, the proximal portion of the shaft has a medial longitudinal depression, but the lower part is flat; a prominent nutrient foramen exists near the articular surface. The dimensions are shown in Table 4.
The genus Megalovis of the Villafranchian are thought by many to be similar to the ancestral form of Ovibos, if not directly ancestral (Crégut-Bonnoure, 1984). Actually, Teilhard de Chardin and Piveteau (1930) had indicated the close affinity of Megalovis piveteaui with Ovibos in the following aspects: cheek teeth without basal pillars, premolars quite hypsodont (Fig. 10A), upper molars with small enamel loops between the lobes at the lingual side (Fig. 10B). Furthermore, the present authors also think the upper molars are quite elongated in crown and the upper premolars with spurs (Fig. 10B). Although the cranial and cornual features are almost completely unknown, the close relationship between Megalovis and ovibovines is verified. On the other hand, the metapodials of Megalovis are also quite stout (Table 4), which resembles those of the recent Ovibos.
Moreover, the SSMZ metacarpal bone has the most similar dimensions with those of M. piveteaui from XSG in Nihewan (Teilhard de Chardin and Piveteau, 1930) except its slightly larger size, but with almost the same stoutness index (Table 4).
The bovid metacarpal bones unearthed at SSMZ site are quite diversified, and most of them are easy to be identified according to their size and proportion, whereas some of them are very similar to each other both in size and stoutness. Therefore, an index should be employed, that’s the robusticity index or stoutness index. Concerning the robusticity index, there exist two calculation methods: one is the relative dimensions of mediolateral diaphysis width and maximum length (SD/GL) ×100) (Maniakas and Kostopoulos, 2017), the other is the distal width/total length (Sher, 1997). Because it’s easier and more practical to measure the distal width, this paper follows Sher’s (1997) method to calculate the robusticity index, i.e. (distal width/greatest length) ×100. Based on the morphological observations and robusticity index calculations, the metacarpal bones of bovids from SSMZ site are quite clearly distinguished (Table 4; Fig. 11).
The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
Megalovis piveteaui and Bison palaeosinensis have quite similar metacarpals both in size and proportion, the only difference is that the formers’ is slightly smaller. Moreover, the Nihewan specimens of M. piveteaui share the similar size and form with M. latifrons of European (Viret, 1954; Radulesco and Samson, 1962) (see Table 4). Megalovis was referred to Ovibovini, maybe its metacarpal is the largest, but less stout, among the ovibovine taxa. M. piveteaui should be larger than Spirocerus wongi in body mass.
Other Images/Table from this Article
-
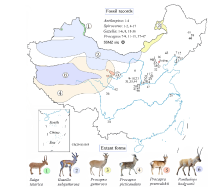 Fig. 1
Location of Shanshenmiaozui (SSMZ) site, with distributional map of living antelopes and related fossil forms of the Pleistocene Epoch in China Data for the extant taxa are from Jiang (2004), Smith and Xie (2008). Fossil localities: 1. Xiashagou of Nihewan; 2. Yushe; 3. Dongdoubi of Yuxian; 4. Jinyuan Cave; 5. Zhoukoudian Loc.1;6. Xujiayao; 7. Mashandong; 8. Salawusu; 9. Xifeng of Qingyang; 10. Loufangzi; 11. Jinniushan;12. Miaohoushan; 13. Haimao of Dalian; 14. Gulongshan; 15. Xiaogushan; 16. Chicheng; 17. Dingcun;18. Xihoudu; 19. Tianshuigou of Dali; 20. Wenxi; 21. Pinglu; 22. Danangou of Yuxian; 23. Bajiazui;24. Gonghe; 25. Yangguo; 26. Tuozidong; 27. Tunliu; 28. Banpo; 29. Linyi; 30. Yuanmou; 31. Heshui;32. Xingtai; 33. Xiaochangcun; 34. Wuwang of Linyi; 35. Gengjiagou; 36. Luhuo; 37. Zhoukoudian Loc.15;38. Upper Cave; 39. Guxiangtun; 40. Zhoujiayoufang; 41. Dali Man site; 42. Aba; 43. Chifeng;44. Rouyuan; 45. Yanjiagang; 46. Yuhongcun of Dali; 47. Lingjing
Fig. 1
Location of Shanshenmiaozui (SSMZ) site, with distributional map of living antelopes and related fossil forms of the Pleistocene Epoch in China Data for the extant taxa are from Jiang (2004), Smith and Xie (2008). Fossil localities: 1. Xiashagou of Nihewan; 2. Yushe; 3. Dongdoubi of Yuxian; 4. Jinyuan Cave; 5. Zhoukoudian Loc.1;6. Xujiayao; 7. Mashandong; 8. Salawusu; 9. Xifeng of Qingyang; 10. Loufangzi; 11. Jinniushan;12. Miaohoushan; 13. Haimao of Dalian; 14. Gulongshan; 15. Xiaogushan; 16. Chicheng; 17. Dingcun;18. Xihoudu; 19. Tianshuigou of Dali; 20. Wenxi; 21. Pinglu; 22. Danangou of Yuxian; 23. Bajiazui;24. Gonghe; 25. Yangguo; 26. Tuozidong; 27. Tunliu; 28. Banpo; 29. Linyi; 30. Yuanmou; 31. Heshui;32. Xingtai; 33. Xiaochangcun; 34. Wuwang of Linyi; 35. Gengjiagou; 36. Luhuo; 37. Zhoukoudian Loc.15;38. Upper Cave; 39. Guxiangtun; 40. Zhoujiayoufang; 41. Dali Man site; 42. Aba; 43. Chifeng;44. Rouyuan; 45. Yanjiagang; 46. Yuhongcun of Dali; 47. Lingjing
-
 Table 1
Studied fossil specimens of small to medium-sized bovids newly unearthed from SSMZ
Table 1
Studied fossil specimens of small to medium-sized bovids newly unearthed from SSMZ
-
 Fig. 2
Partial skulls, jaw bones and teeth of Spirocerus wongi (A-D) and Ovis shantungensis (E) from SSMZ A-D. Spirocerus wongi: A. partial skull with horn-cores (IVPP V 28650), B1-B3. juvenile maxilla with DP2-M1 (V 28651), C1-C4. left mandible with dp2-m1 (V 28652.1), D1-D4. partial right mandible with p4-m3 (V 28653); E1-E2. Ovis shantungensis, maxilla with left DP4-M1 and right DP2-4 and M1 (V 28693) A. anterior view; B1, E1. palatal views; B2, C1, D1, E2. buccal views; C2, D2. lingual views;B3, C3-4, D3-4. occlusal views. The unmarked scale bars equal 20 mm
Fig. 2
Partial skulls, jaw bones and teeth of Spirocerus wongi (A-D) and Ovis shantungensis (E) from SSMZ A-D. Spirocerus wongi: A. partial skull with horn-cores (IVPP V 28650), B1-B3. juvenile maxilla with DP2-M1 (V 28651), C1-C4. left mandible with dp2-m1 (V 28652.1), D1-D4. partial right mandible with p4-m3 (V 28653); E1-E2. Ovis shantungensis, maxilla with left DP4-M1 and right DP2-4 and M1 (V 28693) A. anterior view; B1, E1. palatal views; B2, C1, D1, E2. buccal views; C2, D2. lingual views;B3, C3-4, D3-4. occlusal views. The unmarked scale bars equal 20 mm
-
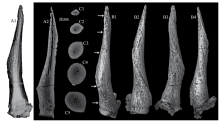 Fig. 3
CT scan images and 3-D reconstructions of the horn-core of Spirocerus wongi from SSMZ (IVPP V 28650) A1-A2. CT scan images showing the general canal system (A1) and a longitudinal slice (A2);C1-C5. CT scan slices showing the changes of cross sections and the canal system at different levels;B1-B4. CT image reconstruction of the right horn-core in anterior (B1), lateral (B2), posterior (B3) and medial (B4) views
Fig. 3
CT scan images and 3-D reconstructions of the horn-core of Spirocerus wongi from SSMZ (IVPP V 28650) A1-A2. CT scan images showing the general canal system (A1) and a longitudinal slice (A2);C1-C5. CT scan slices showing the changes of cross sections and the canal system at different levels;B1-B4. CT image reconstruction of the right horn-core in anterior (B1), lateral (B2), posterior (B3) and medial (B4) views
-
Table 2
Measurements of the horn-cores of Spirocerus wongi, compared with related species (mm)
-
Table 3
Measurements of the teeth of Spirocerus wongi, compared with related species (mm)
-
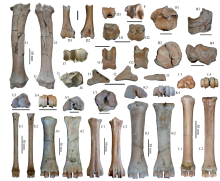 Fig. 4
Postcranial bones of small and medium-sized bovids from SSMZ A-C. Spirocerus wongi: A1-A3. left Mc III+IV (IVPP V 28655),B1-B4. right naviculo-cuboid+lat-mid+medial cuneiforms (V 28656.2-3),C1-C3. right Mt III+IV (V 28656.4); D-I. Gazella sinensis: D1-D3. partial left humerus (V 28687),E1-E4. left Mc III+IV (V 28688), F. distal epiphysis of left tibia (V 28691),G1-G2. left astragalus (V 28692), H1-H2. left naviculo-cuboid (V 28690), I1-I3. 3rd phalanx (V 28689);J-K. Megalovis piveteaui: J1-J4. left radius (V 28654), K1-K4. right Mc III+IV (V 28695);L1-L4. Ovis shantungensis: left Mc III+IV (V 28694). A1, C1, D1, E1, G1, J1, K1, L1. anterior views;A2, B4, C2, D2, E2, G2, J2, K2, L2. posterior views; A3, B1, C3, E3, H1, J3, K3, L3. proximal views;B2, D3, E4, F, H2, J4, K4, L4. distal views; B3. medial view; I1. lateral view; I2. interdigital view;I3. volar view. The arrows indicate the lateral projection of the radius. The unmarked scale bars equal 20 mm
Fig. 4
Postcranial bones of small and medium-sized bovids from SSMZ A-C. Spirocerus wongi: A1-A3. left Mc III+IV (IVPP V 28655),B1-B4. right naviculo-cuboid+lat-mid+medial cuneiforms (V 28656.2-3),C1-C3. right Mt III+IV (V 28656.4); D-I. Gazella sinensis: D1-D3. partial left humerus (V 28687),E1-E4. left Mc III+IV (V 28688), F. distal epiphysis of left tibia (V 28691),G1-G2. left astragalus (V 28692), H1-H2. left naviculo-cuboid (V 28690), I1-I3. 3rd phalanx (V 28689);J-K. Megalovis piveteaui: J1-J4. left radius (V 28654), K1-K4. right Mc III+IV (V 28695);L1-L4. Ovis shantungensis: left Mc III+IV (V 28694). A1, C1, D1, E1, G1, J1, K1, L1. anterior views;A2, B4, C2, D2, E2, G2, J2, K2, L2. posterior views; A3, B1, C3, E3, H1, J3, K3, L3. proximal views;B2, D3, E4, F, H2, J4, K4, L4. distal views; B3. medial view; I1. lateral view; I2. interdigital view;I3. volar view. The arrows indicate the lateral projection of the radius. The unmarked scale bars equal 20 mm
-
 Fig. 5
Incomplete skulls and horn-cores of Gazella sinensis from SSMZ A. partial skull of a juvenile with horn-cores (IVPP V 28658); B1-B3. partial skull with horn-cores (V 28657); C1-C2. left and right horn-cores (V 28659.1, V 28659.2); D1-D2. partial skull with left horn-core (V 28667);E. partial skull with right horn-core (V 28661). A, B1, C1-2, D1. anterior views; B3. posterior view;B2. dorsal view; E. medial view; D2. lateral view. Scale bars equal 50 mm
Fig. 5
Incomplete skulls and horn-cores of Gazella sinensis from SSMZ A. partial skull of a juvenile with horn-cores (IVPP V 28658); B1-B3. partial skull with horn-cores (V 28657); C1-C2. left and right horn-cores (V 28659.1, V 28659.2); D1-D2. partial skull with left horn-core (V 28667);E. partial skull with right horn-core (V 28661). A, B1, C1-2, D1. anterior views; B3. posterior view;B2. dorsal view; E. medial view; D2. lateral view. Scale bars equal 50 mm
-
Table 5
Measurements of partial cranial bones and horn-cores of Gazella sinensis (mm)
-
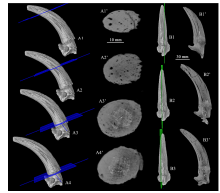 Fig. 6
CT scan images of the horn-core of Gazella sinensis (IVPP V 28661) from SSMZ A1-A4. CT image reconstruction showing positions of the cross CT scan slices (A1’-A4’);B1-B3. CT image reconstruction showing positions of the longitudinal CT scan slices (B1’-B3’)
Fig. 6
CT scan images of the horn-core of Gazella sinensis (IVPP V 28661) from SSMZ A1-A4. CT image reconstruction showing positions of the cross CT scan slices (A1’-A4’);B1-B3. CT image reconstruction showing positions of the longitudinal CT scan slices (B1’-B3’)
-
 Fig. 7
Jaw bones and teeth of Gazella sinensis from SSMZ (A-J), compared with related taxa (K-P) A. maxilla with P3-M1 (IVPP V 28673); B. maxilla with M1-3 (V 28674);C1-C4. left mandible with p2-m3 (V 28680); D1-D3. left mandible with p2-m3 (V 28681.2);E1-E2. left mandible with dp2-4 and m1-2 (V 28685.1), E3. detail of dp2-4; F. right p4-m3 (V 28677);G. left p3-m3 (V 28682); H. right p2-m3 (V 28675); I. left p4-m3 (V 28686); J. left p4-m3 (V 28678);K. Procapra przewalskii, right p2-m3 (horizontally flipped) (NWIPB-0001172♀);L. Procapra gutturosa, left p2-m3 (NWIPB 620032); M. Gazella subgutturosa, left p2-m3 (NWIPB 609001);N. Pseudois nahaur, left p2-m3 (NWIPB-KX1); O. Capra ibex, left p2-m3 (IOZ-2);P. Ovis ammon, left p2-m3 (OV 1346-2). A, B, C1, C4, D1, E3, F-P. occlusal views;C2, D2, E1. buccal views; C3, D3, E2. lingual views. The arrows indicate the variations of p4 The unmarked scale bars equal 20 mm
Fig. 7
Jaw bones and teeth of Gazella sinensis from SSMZ (A-J), compared with related taxa (K-P) A. maxilla with P3-M1 (IVPP V 28673); B. maxilla with M1-3 (V 28674);C1-C4. left mandible with p2-m3 (V 28680); D1-D3. left mandible with p2-m3 (V 28681.2);E1-E2. left mandible with dp2-4 and m1-2 (V 28685.1), E3. detail of dp2-4; F. right p4-m3 (V 28677);G. left p3-m3 (V 28682); H. right p2-m3 (V 28675); I. left p4-m3 (V 28686); J. left p4-m3 (V 28678);K. Procapra przewalskii, right p2-m3 (horizontally flipped) (NWIPB-0001172♀);L. Procapra gutturosa, left p2-m3 (NWIPB 620032); M. Gazella subgutturosa, left p2-m3 (NWIPB 609001);N. Pseudois nahaur, left p2-m3 (NWIPB-KX1); O. Capra ibex, left p2-m3 (IOZ-2);P. Ovis ammon, left p2-m3 (OV 1346-2). A, B, C1, C4, D1, E3, F-P. occlusal views;C2, D2, E1. buccal views; C3, D3, E2. lingual views. The arrows indicate the variations of p4 The unmarked scale bars equal 20 mm
-
Table 6
Measurements of teeth of Gazella sinensis from SSMZ, compared with those from Xiashagou (mm)
-
 Fig. 8
Comparison of premolar series among some Quaternary gazelle species of China A. Gazella sinensis, left p2-4, IVPP V 28681.2, SSMZ; B. Procapra gutturosa, left p2-4, NWIPB 0006065, extant; C. P. przewalskii, right p2-4 (horizontally flipped), NWIPB 0001172, extant;D. P. picticaudata, left p2-4, NWIPB 0001179, extant; E. Gazella subgutturosa, left p2-4, IVPP-c-05, extant;F. Saiga tatarica, right p3-4 (horizontally flipped), NWIPB S-80503, extant;G. Pantholops hodgsonii, left p3-4, NWIPB 77001, extant. All are in occlusal views
Fig. 8
Comparison of premolar series among some Quaternary gazelle species of China A. Gazella sinensis, left p2-4, IVPP V 28681.2, SSMZ; B. Procapra gutturosa, left p2-4, NWIPB 0006065, extant; C. P. przewalskii, right p2-4 (horizontally flipped), NWIPB 0001172, extant;D. P. picticaudata, left p2-4, NWIPB 0001179, extant; E. Gazella subgutturosa, left p2-4, IVPP-c-05, extant;F. Saiga tatarica, right p3-4 (horizontally flipped), NWIPB S-80503, extant;G. Pantholops hodgsonii, left p3-4, NWIPB 77001, extant. All are in occlusal views
-
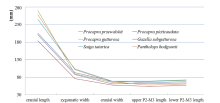 Fig. 9
Toothrow length and cranial size of extant gazelles in China Raw data was employed from Jiang, 2004
Fig. 9
Toothrow length and cranial size of extant gazelles in China Raw data was employed from Jiang, 2004
-
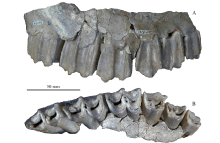 Fig. 10
Maxilla of Megalovis piveteaui (MNHN-NIH 150) from Xiashagou of Nihewan A. buccal view; B. occlusal view
Fig. 10
Maxilla of Megalovis piveteaui (MNHN-NIH 150) from Xiashagou of Nihewan A. buccal view; B. occlusal view
-
 Fig. 11
Length vs distal width of metacarpal bones of diverse bovids The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
Fig. 11
Length vs distal width of metacarpal bones of diverse bovids The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
|
